|
|
|
L’anatomie et les fonctions des cellules
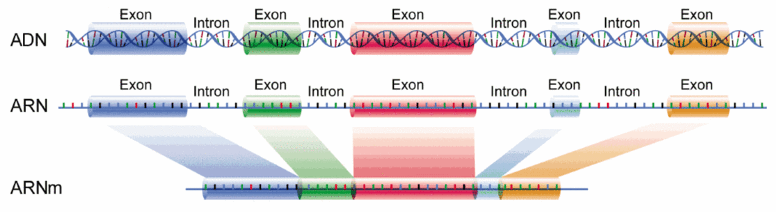
Le deuxième code génétique (III) Depuis les années 1980, les généticiens ont remarqué que le code génétique ne suffisait pas à expliquer la diversité des protéines (des tissus biologiques) : un gène peut conduire à la fabrication de plusieurs protéines différentes, la cellule étant capable de produire des milliers d'ARN messagers (ARNm). Inversement, les généticiens ne retrouvaient pas le texte exact de l'ADN à l'origine d'une protéine. Il leur manquait donc une information, comme si durant le processus de l'épissage (le séquençage de l'ADN lors de la transcription de l'ADN en ARN) il existait un code intermédiaire définissant quel type d'ARN messager devait être produit en fonction de la nature du gène ou du tissu concerné. C'est ce qu'ont découvert en 2010 Benjamin Blencowe et Brendan Frey de l'Université de Toronto en perçant le mystère entourant l'épissage alternatif (alternative splicing). Ou comment, à partir de seulement 26500 gènes exprimés, le génome humain génère des informations génétiques bien plus nombreuses et complexes, contrôlant la plupart des activités cellulaires. Une étude publiée dans la revue "PLoS One" en 2017, Helmut Schiessel de l'Université de Leyde et son équipe ont confirmé l'intervention de ce deuxième code génétique dans le pliage des molécules et son impact possible sur les mutations génétiques. Grâce à ce deuxième code génétique qui permet de décupler le potentiel de l'ADN, l'ARN prémessager est découpé puis réassemblé selon le code de l'épissage. L'ARN est ensuite décodé et l'ARN messager est constitué. Concrètement, les bases nucléiques doivent se recombiner par paires et de façon unique : A avec T et G avec C. Lorsque les nucléotides des ARN trouvent leur base complémentaire, une enzyme polymérase les assemble avec l’ARN messager. Cette transcription suit donc le même ordre que celui des séquences de l’ADN et peut se répéter un très grand nombre de fois. Les séquences d’ADN qui sont transcrites mais non traduites en protéines sont dénommées introns (cf. S.M. Berget et al., 1977). Elles sont détruites lors de l'épissage. Celles qui sont traduites sous forme de protéines et véhiculées par l’ARN messager sont dénommées exons. Mis bout-à-bout, les exons forment une séquence qui varie d'un tissu à l'autre. Au terme de cette étape, l'ARN prémessager donne naissance à un ARN messager spécifique à chaque tissu. Les exons sont ensuite exprimés pour fabriquer une protéine. Au cours de la transcription de l’ADN, les gènes assurent plusieurs tâches : avant toute chose ils synthétisent des ARN fonctionnels, surtout des ARN de transfert (ARNt) dont certains sont débités et mis à dimension par des enzymes contenant des ARN à fonction catalytique (telle la ribonucléase P chez le fameux colibacille Escherichia coli). Il faut insister sur ce point car cette molécule d’ARN est capable de catalyser une réaction presque aussi efficacement que le complexe réplisome ! La transcription de l’ADN sert également à synthétiser des ARN ribosomiaux (ARNr) qui assurent la croissance des bactéries (transcription des gènes). A
voir : Modélisation
de la formation de la chromatine et des nucléosomes Processus de pliage de l'ADN pour former des protéines de chromatine dans le noyau cellulaire A lire : Le deuxième code génétique (Pour La Science, 2010) Découverte du code de l'épissage alternatif
L'ADN non exprimé Ainsi que nous l'avons expliqué en première page, les deux génomes procaryotes et eucaryotes se différencient également par la présence chez les cellules eucaryotes d’une beaucoup plus grande proportion d’ADN porteur de gènes non fonctionnels. En effet, à côté des quelques 26500 gènes soit à peine 2% du génomène humain lourds de sens codant des protéines cellulaires, plus de 97% ne sont pas exprimés ! Il s’agit de fragments d’ADN contenant des séquences régulatrices, des traces de duplications et de mutations antérieures, des gènes, des virus et des rétrovirus morts ou inactifs que l’évolution n’a pas éliminé. La présence de cette "pourriture" (junk)[4] comme l’a si bien considérée Crick et que certains biologistes appellent l'ADN "poubelle" est longtemps restée une énigme et considérée comme inutile. En fait, les généticiens s'étaient en partie trompés. Toutefois, les chercheurs se doutaient que ces séquences non exprimées devaient probablement contenir des séquences connectrices, comprenant divers signaux régulateurs, des gènes structuraux et dupliqués, et la probabilité que chaque gène soit précédé d’une région importante utilisée par la machinerie lors de la transcription. Il fallut une génération et attendre 2019 pour qu'on dispose enfin du séquençage complet du génome humain et qu'on y voit un peu plus clair. Ce génome déchiffré intégralement est nommé T2T-CHM13 (cf. S.Nurk et al., 2021; Nature, 2021; S.Nurk et al., 2022). Les transposons ou gènes sauteurs Si le bon sens nous faire dire que ce serait une perte de temps et d'énergie pour la cellule que de trier et de gérer "durablement" l'énorme quantité d'ADN de rebut, il faut bien reconnaître que continuer inlassablement à les dupliquer sans que personne ne se souvienne pourquoi, est tout aussi inutile et est un gaspillage d'énergie très précieuse. Étrange stratégie même paradoxale de la part d'un organisme par ailleurs excellement bien organisé. Mais notre compréhension du fonctionnement du génome n'est peut-être pas aussi claire et complète qu'on l'imagine. En fait, on a pu résoudre une partie de cette énigme. Et il a bien fallu reconnaître que les conclusions initiales des généticiens étaient prématurées. Comme le disait le dialoguiste Michel Audiart "C'est pas parce qu'on n'a rien à dire qu'il faut fermer sa gueule", expression reprise dans le film éponyme réalisé par Jacques Besnard en 1975. Les gènes ont un peu la même mentalité, en particulier certains gènes non exprimés qui veulent se rendre utiles.
Dans les années 1950, la cytogénéticienne Barbara McClintock (1902-1992) découvrit les "gènes sauteurs" ou transposons dans l'ADN de maïs pour laquelle elle reçut le prix Nobel de Physiologie (Médecine) en 1983, le temps que les généticiens compennent la portée de ses travaux précuseurs (McClintock arrêta de publier ses travaux en 1953 car ses théories n'étaient ni comprises ni acceptées par ses pairs jusqu'à ce que les généticiens s'en rappellent dans les années 1960-1970). Les généticiens ont constaté que le génome et pas seulement humain n'est pas une structure passive mais que des séquences d'ADN peuvent se détacher, s'autorépliquer et s'insérer en d'autres endroits, créant apparemment un désordre indescriptible que seule la machinerie cellulaire est capable de décrypter. Ces séquence de gènes sauteurs sont les transposons de McClintock. Si aujourd'hui on considère qu'ils jouent un rôle essentiel dans la stucture et le fonctionnement du génome, initialement et par manque d'outils adaptés à leur étude, ils furent considérés comme des gènes parasites ou des gènes égoïstes (lors de la sélection naturelle, la sélection des gènes l'emporte sur la sélection des organismes et des populations, cf. le livre "Le gène égoïste" de Richard Dawkins), voire pire, de l'ADN de rebut juste bon pour la poubelle ! Or aujourd'hui, les transposons sont considérés comme des éléments actifs essentiels susceptibles d'augmenter le potentiel évolutif des organismes. Portant eux-mêmes des séquences régulatrices, ils interviennent par exemple au cours des mutations, de la recombinaison, dans les séquences codantes ou régulatrices de gènes (quitte à créer des interférences), bref tout au long de l'évolution du génome. Ils sont capables de remodeler la structure chromosomique afin que les gènes s'expriment différemment et de faire émerger de nouvelles fonctions génétiques de grande ampleur. Les transposons interviennent notamment dans les phénomènes épigénétiques (voir plus bas). Parmi les découvertes que l’on espère de la cartographie du génome humain qui fut terminée en 2001 réside l’espoir que la "pourriture" ne codant pas pour les protéines recèle un enregistrement de l’histoire de l’évolution humaine. En effet, si nous pouvions identifier l'origine et le rôle éventuel de ces séquences inactives, les chercheurs pourraient sans doute mieux comprendre notre évolution, y compris certains de nos comportements que les réactions biochimiques seules ne peuvent totalement expliquer (comme par exemple certaines actions inconscientes, l'attirance sexuelle, etc.). Ainsi, en 2005 des chercheurs ont découvert dans ces ADN non exprimés des séquences dérivées de gènes et des traces de virus et d'antivirus ayant participé aux mutations génétiques puis écartés au cours de l’évolution. Parmi ceux-ci on retrouve les transposons qui ont été répertoriés en 3 types regroupés en 2 classes : la classe I des rétrotransposons et la classe II des transposons à ADN qui se différencient par leur structure et leurs mécanismes de transposition. Ils comprennent entre 1000 et 20000 nucléotides capables de coder les protéines dont les enzymes nécessaires à leur fonction. Ces transposons contiennent eux-mêmes des sous-unités telles que les MITE (Miniature Inverted-repeat Transposable Element), les LINE et les SINE (Long et Short Interspersed Nuclear Elements) d'une taille variant entre 100 et 1000 nucléotides qui sont clairement des traces de rétrovirus endogènes humains ou HERV. Ils ne sont pas autonomes et dépourvus de séquence codante et sont obligés d'utiliser des protéines codées par d'autres transposons. Nous verrons à propos des cas de recontamination par le Covid-19 que le rétrotransposon LINE1 a permis d'intégrer une séquence génétique du virus dans le génome humain. mais ce n'est pas la seule trace virale que possèdent nos cellules. Les transposons représentent environ 20% du génome humain (cf. R.Cordaux et M.Batze, 2009) et ont construit environ 40% de notre génome (cf. J.L. Goodier, 2016). Du coup, les scientifiques ne voient plus les virus sous le même angle et certains y verraient même des alliés. Nous reviendrons sur le rôle des virus dans l'évolution. En 2012, dans le cadre du projet ENCODE, des scientifiques ont découvert que 75% de l'ADN non codant du génome humain participaient à la transcription et presque 50% (contre 90% dans le génome du blé) étaient ainsi mis à disposition des protéines impliquées dans le contrôle génétique. Le rôle de l'ADN ultraconservé En 2012, il était encore hasardeux de conclure que ces séquences non codantes intervenaient dans les facteurs de transcription. Mais en réalisant de nouvelles expériences, au fil des années les généticiens ont constaté que cet ADN non codant joue un rôle biochimique et que la transciption de ces segments est nécessaire mais on ne peut pas encore affirmer qu'ils offrent un quelconque avantage en terme d'évolution si ce n'est que certaines séquences assurent la pérénité de l'espèce. C'est notamment le cas de l'ADN ultraconservé.
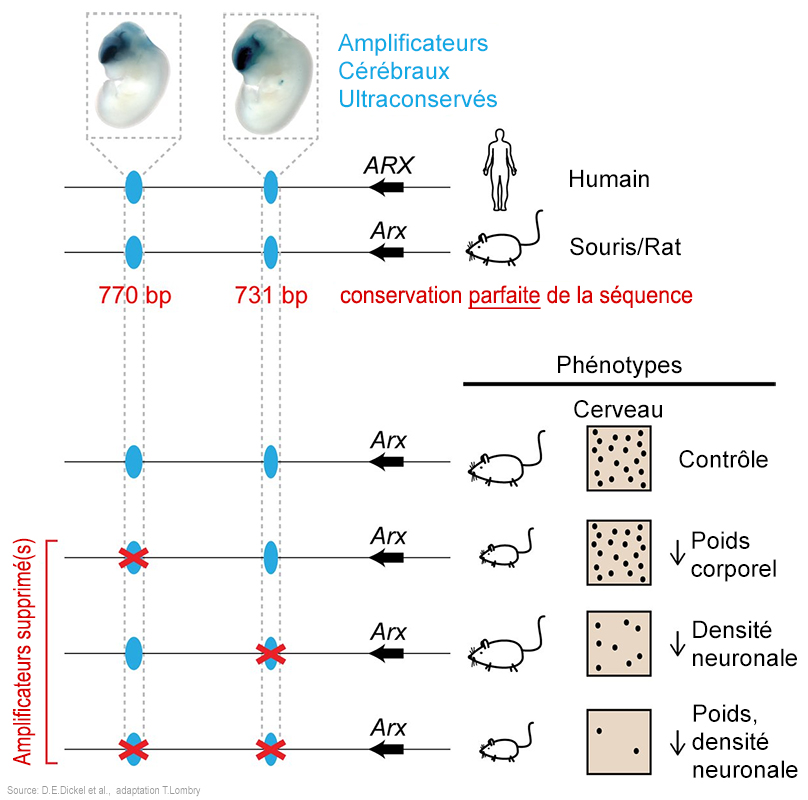
Les généticiens ont trouvé dans le génome de la souris au moins 200 paires de nucléotides intégrées dans des séquences d'ADN non codant qui sont restées intactes depuis environ 80 millions d'années, ce qu'on appelle des séquences ultraconservées, mais qui ne sont pas inutiles pour autant. Parmi celles-ci il y a quatre séquences situées sur le chromosome sexuel X, à proximité du gène Arx (un gène affectant le développement cérébral et dont la mutation est associée chez l'homme à l'autisme) qui amplifient l'expression de ce gène. Des chercheurs ont voulu savoir dans quelle mesure ces séquences ultraconservées étaient encore utilisées par la machinerie génomique. En 2018, une équipe de chercheurs dirigée par Diane E. Dickel du Laboratoire Lawrence-Berkeley annonça dans la revue "Cell" que la suppression de ces ADN non codant par une technique d'édition génomique appelée CRISPR-Cas9 n'affecte pas la viabilité des souris mais altère néanmoins leur développement. La suppression de l'un des quatre amplificateurs ou des combinaisons de deux d’entre eux produit un retard de croissance du foetus tandis que la suppression de deux amplificateurs produit une anomalie du développement cérébral. Plus étrange, l'altération n'est pratiquement pas détectable in vitro mais uniquement in vivo. Enfin, si on supprime les quatre séquences amplificatrices, l'animal devient stérile. Autrement dit, au cours de l’évolution, sous la pression de sélection la souris a conservé des séquences non codantes mais dont les fonctions demeurent essentielles; leur modification s'avère fatale ou conduit à la stérilité de l'animal. Au vu de cette expérience, on ne peut donc pas affirmer que la suppression de séquences ultraconservées n'a pas d'impact sur l'organisme. Ceci dit la recherche continue pour déterminer si d'autres séquences ont une importance similaire dans le génome humain. Les généticiens fondent beaucoup d'espoirs dans le séquençage du génome à la fois dans le but de faire avancer la thérapie génique mais également afin de comprendre la complexité de l'évolution et la nature humaine. Notons qu'à l’exception de quelques gènes d’ARN de transfert et d’ARN ribosomial, ces ADN de rebut n’existent pratiquement pas chez les cellules procaryotes. La raison la plus probable est que le génome bactérien a été "amélioré" par la sélection naturelle afin d’assurer une multiplication aussi rapide que possible. Ainsi, dans les bactéries en croissance, la réplication de l’ADN ne cesse pratiquement jamais. Les phénomènes épigénétiques Parmi les nombreuses questions ouvertes de la génétique, il y a celle des phénomènes dits épigénétiques. Suite à un stress, généralement d'origine environnementale, le génome de l'organisme produit des modifications secondaires transmissibles et réversibles mais sans effet sur la séquence des nucléotides (à l'exception des cancers ou du vieillissement où ils interviennent également). Cela permet par exemple à une espèce de s'adapter au climat des basses terres ou au climat d'altitude en modifiant en quelques générations (quelques décennies chez les fleurs, quelques milliers d'années ches les humains) toute sa structure (par exemple sa taille, ses performances, sa couleur, etc). Même un astronaute subit des modifications épigénétiques lors d'un séjour prolongé dans l'espace (cf. l'étude conduite sur Scott Kelly de la NASA). Les transposons précités interviennent dans les phénomènes épigénétiques en favorisant les mutations. La théorie de l'évolution explique que les organismes vivants survivent et les espèces se transforment au cours du temps grâce à l'expression des gènes et au hasard de la sélection naturelle associé aux mutations aléatoires. Mais il saute aux yeux qu'au sein d'un même règne, tous les animaux, comme tous les insectes ou tous les végétaux présentent globalement les mêmes formes. Charles Darwin et D'Arcy Thompson avaient déjà constaté que les différences entre les espèces pouvaient s'expliquer par des différentes morphologiques. En prenant pour exemple la forme du bec des pinsons des Galápagos, Darwin constata que leur bec évoluait progressivement d'une espèce à l'autre suite à des mutations et la sélection naturelle, fondant la théorie de l'évolution. A son tour d'Arcy Thompson découvrit que la forme des poissons obéissait à des lois de transformations. Ainsi, récemment on a découvert que l'homothétie agit sur la taille et la transvection agit sur l'inclinaison des formes. Ces deux facteurs géométriques combinés permettent d'expliquer la diversité des formes des espèces. Ces transformations seraient contrôlées par des "gènes architectes" et notamment par la protéine Bmp4. Le rôle des gènes Hox Le génome n'est pas un grand livre qu'il suffit de lire pour construire une cellule ou un organisme. Ce n'est pas un plan de construction au sens strict mais plutôt une base de données qui indique comment utiliser les briques élémentaires de l'ADN pour créer un organisme viable. Au stade embryonnaire de la gastrula (14-15e jour de l'embryogenèse humaine), l'embryon ressemble encore à une petite sphère de mosaïques contenant déjà plusieurs milliers de cellules qui s'organisent pour former les bases du plan corporel. Une invagination apparaît qui va former trois feuillets ou couches de cellules (ectoderme, mésoderme et endoderme). A ce stade, on constate que l'animal en devenir a le choix entre deux évolutions : certaines espèces animales sont protostomiens (la bouche se forme avant l'anus) comme les annélidés et les calmars, d'autres sont deutérostomiens (l'anus se forme en premier) comme les êtres humains, les oursins ou les étoiles de mer. Si ce classement qui remonte aux travaux de Karl Grobben en 1908 n'est plus très représentatif de l'évolution et bien que sa pertinence fasse débat (cf. Intermédiairepub), il montre que la Nature a des contraintes dès la création des individus et n'est pas totalement aveugle comme certains évolutionnistes aimeraient le croire. Ainsi si on retire une cellule au stade 4 cellules de l'oeuf (l'oeuf contient 4 cellules de même volume) d'un protostomien, son évolution est interrompue car il est incapable de se développer et meurt. En revanche, en effectuant la même manipulation au même stade, les cellules des deutérostomiens peuvent encore être reprogrammées et ce jusqu'à un stade avancé de leur développement. Ensuite, on observe une segmentation de l'embryon le long de l'axe antéro-postérieur (cf. T.F. Schilling et R. D. Knight, 2001; G.Balavoine et al., 2010; M. Diaz-Cuadros et al., 2021), puis tout se forme à peu près en même temps mais dans un ordre bien précis selon l'expression des gènes.
Après la découverte des gènes à la fin du XIXe siècle et l'effet des mutations, puis de l'ADN et déchiffré le code génétique (voir page 1), en 1983 des généticiens ont découvert une catégorie particulière de gènes homéotiques, c'est-à-dire de gènes qui déterminent le plan corporel, appelés les gènes Hox. Hox est l'abréviation du terme anglais Homeobox (homéoboîte) par référence à la séquence ADN qui, après traduction, code une séquence d'acides aminés (un homéodomaine) dans les protéines. Les gènes Hox jouent un rôle plus important que les autres dans le développement embryonnaire. En effet, les gènes Hox sont capables de déléguer des actions à d'autres gènes et même d'activer d'autres gènes Hox pour terminer la formation de l'extrémité ou de la jointure d'un membre par exemple. On peut considérer les gènes Hox comme des architectes, tandis que tous les autres gènes sont les ouvriers qui construisent l'édifice grâce aux briques et au ciment que sont les protéines qu'ils produisent.
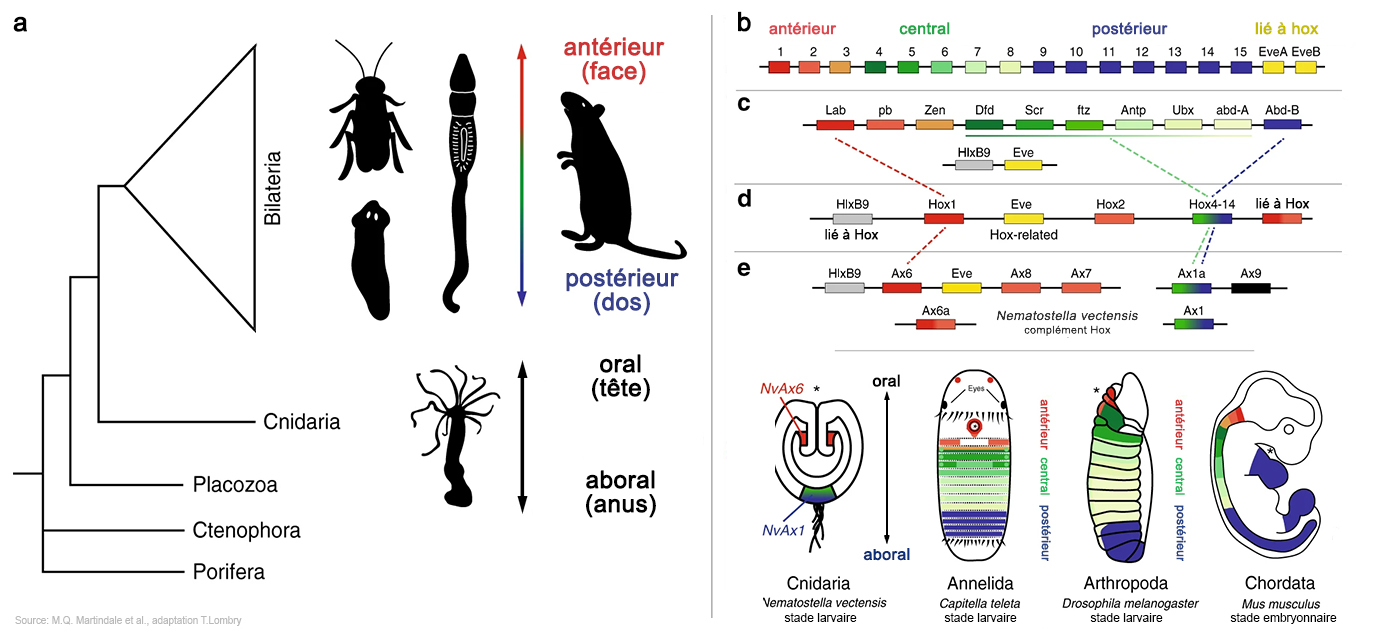
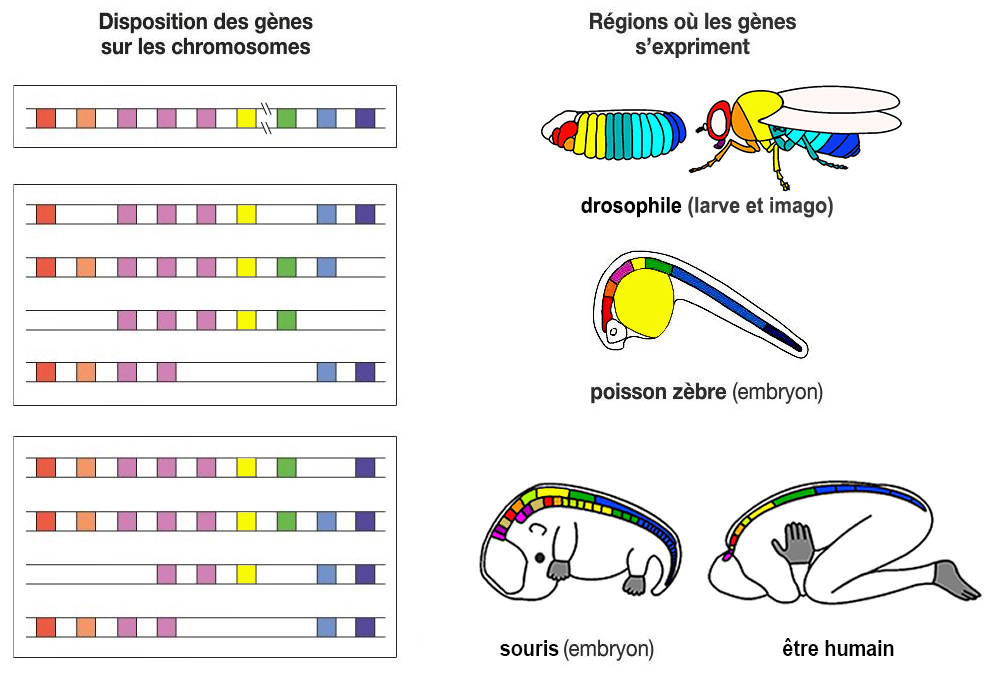
Mais comme les architectes, les gènes Hox ne contrôlent pas le résultat. A défaut d'entrepreneur général ou d'auditeur, ce sont les gènes qui s'assurent de produire suffisamment de matière première pour fabriquer la partie de l'organisme qui les concerne mais leur pouvoir de contrôle est également limité, d'autant moins qu'ils peuvent eux-mêmes être victimes de mutations à l'origine de malformations ou de cancers. Il existe des gènes Hox chez tous les animaux bilatériens (à symétrie bilatérale, dont les deux côtés sont identiques). Mais si nous sommes construits à partir des mêmes gènes, pourquoi la grenouille ne ressemble pas au chat et l'être humain à un papillon ? Simplement parce que si nous avons les mêmes gènes architectes Hox, ils activent des gènes propres à chaque espèce. Pourquoi les animaux ont une tête et un anus, des côtés antérieur (oral) et postérieur (aboral) - on parle de face et de dos chez les humains - et des membres attachés au thorax et à l'abdomen (en gros) ? Deux découvertes expliquent cette disposition anatomique. On a découvert que les gènes Hox de la mouche et de la souris par exemple sont disposés dans un ordre précis. La mouche possède 8 gènes Hox alignés sur un seul chromosome, formant un cluster de gènes nommé le complexe de gènes Hox (HOX-C). Alors que beaucoup d'animaux possèdent beaucoup plus ou beaucoup moins de chromosomes que l'être humain, la souris et tous les autres mamifères possèdent 13 gènes Hox également bien alignés mais dispersés sur 4 chromosomes (chr. 7, 17, 12 et 2 chez l'être humain) plus quelques doublons (cf. P.W. Holland et al., 2007; P.Z. Myers, 2008; A.Durston et al., 2012). On a également découvert qu'au cours du développement, les gènes Hox s'activent séquentiellement, selon leur position sur le chromosome : Hox1, puis Hox 2, Hox 3, etc. Hox1 code pour la région postérieure de la tête, Hox2 et Hox3 pour le haut du cou, Hox4 pour la base du cou, Hox5 pour les épaules (la ceinture scapulaire), etc. jusqu'au thorax, l'abdomen et la queue. Chaque gène Hox va ensuite activer en cascade toute une série d'autres gènes dans les régions concernées de l'organisme en développement de façon à ce que par exemple les yeux soient bien placés dans la tête et que la main comprenne bien 5 doigts séparés. En résumé, l'architecture du corps repose sur trois organisations parallèles : - L'ordre des gènes sur les chromosomes - L'ordre d'activation temporelle - L'ordre sur le corps de l'animal. Chaque gène Hox définit l'architecte d'une partie de l'organisme, l'ensemble des gènes Hox définissant le schéma de construction de l'organisme (où est l'avant avant, où est l'arrière, la succession des "modules" d'avant en arrière et leur emplacement dans le corps), comme un architecte définit les modules composant un édifice et l'ordre de construction. Chaque Hox va ensuite s'occuper d'activer les gènes spécifiques de son "module" qui, comme les ouvriers feront le nécessaire pour construire l'édifice sans avoir besoin d'un plan général de construction. Comme des boîtes de Lego ou de Meccano composées de différents pièces élémentaires nous permettent de construire des objets fonctionnels, ainsi procède dame Nature avec les gènes. A consulter : Homeobox Database Base de données des gènes homéotiques
Mais cette organisation à la fois séquentielle et parallèle n'explique pas tout. Par exemple, comment sont conçues les jointures, comment les "modules" des parties du corps sont assemblés entre eux pour obtenir un corps final complet et fonctionnel ? Concrètement pourquoi les bras sont fixés aux épaules et pas ailleurs sur le tronc ? Dans le cas des bras, c'est le gène Hox5 qui s'occupe de la ceinture scapulaire et uniquement de celle-ci. Il doit donc faire appel à d'autres gènes pour attacher correctement le bras droit à l'épaule droite. Etant donné que le dernier Hox de la série n'est pas encore intervenu, Hox5 fait appel à Hox13. Ce dernier crée deux copies d'Hox13 et de son cluster de gènes ouvriers qui vont assembler le bras au bon endroit. Cependant parfois le gène Hox13 est à l'origine de malformations des membres (polydactylie et atrophie). Autre constat, pourquoi nos mains ne ressemblent pas à des pattes de chat (même si certains parviennent à écrire de cette façon !) ? Simplement parce que si les gènes élémentaires sont composés des mêmes nucléotides, ils sont différents chez chaque espèce. Comme le puzzle d'un être humain est différent du puzzle d'un chat, les génomes des deux espèces ne sont pas compatibles et les pièces de l'un ne s'adaptent pas à l'autre. Les gènes Hox existent depuis 600 millions d'années et l'ancêtre commun des vertébrés, des arthropodes et des annélidés (juste avant l'explosion du Cambrien). Ils ont été conservés car leur rôle est primordial dans l'édification des organismes, le moindre changement déclenchant des modifications anatomiques importantes. Y toucher par génie génétique reviendrait à jouer à Frankenstein. On reviendra sur les manipulations génétiques. Le déterminisme génétique et le rôle de l'éducation Si le génome représente la mémoire avec laquelle un organisme s'adapte et perdure dans son environnement, aujourd'hui les chercheurs (biologistes moléculaires, experts des neurosciences, psychologues, psychiatres) reconnaissent que l'ADN d'un individu n'est pas le seul facteur caractérisant son caractère et son intelligence; les effets induits par les contraintes physiques et l'éducation vont également déterminer son évolution.
En 2018, une équipe de chercheurs dirigée par Ian J. Deary de l'Université d'Edimbourg annonça dans la revue "Molecular Psychiatry" qu'ils avaient identifié 538 gènes liés à l'intelligence dans 187 loci (régions du génome) associés aux facultés intellectuelles. Mais cela ne signifie pas que toute notre intelligence dépend de ce potentiel génétique. Des psychiatres ont démontré après plusieurs années d'analyses cliniques de patients, qu'un homme génétiquement classé comme psychopathe et dont le cerveau fonctionnait comme tel (les PET Scans montrent une faible activité des aires préfrontales et pariétales, comme c'est généralement le cas chez les criminels) peut évoluer dans le bon sens et devenir "pro-social", c'est-à-dire une personne très sociable, gentille et même créative et ayant de l'empathie pour ses semblables. Ce fut notamment le cas du neuroscientifique James Fallon (cf. aussi ce reportage d'ARTE). Cela signifie que bien que porteur d'un bagage génétique présentant des gènes dont les allèles sont faibles (par ex. MAO-A lié à l'agressivité) et donc à risque, un enfant va bien ou "mal tourner" en fonction de l'éducation qu'il recevra, autrement dit de la pression de l'environnement combinée à ses facultés intellectuelles et donc ses prédispositions génétiques. Cela signifie aussi que le génome en soi n'est ni bon ni mauvais. Tout va dépendre de ses interactions avec l'environnement et de la manière dont son porteur va réagir en fonction des évènements. Comme le titrait le "Time" en 2010, voilà "pourquoi l'ADN n'est pas ton destin". Selon une étude publiée dans la revue "Current Biology" en 2013 par Ian J. Deary, le génome ne contribue qu'à environ 50% de l'intelligence d'une personne, une valeur plus faible chez l'enfant (40% à 9 ans) que chez l'adulte (70-80%). Bref, une bonne partie des aptitudes et des stimulations intellectuelles dépendent de l'environnement. On reviendra sur les facteurs déterminant l'intelligence dans l'article consacré au système nerveux. CRISPR : les espoirs et les risques des manipulations génétiques Les deux principales méthodes utilisées par les organismes multicellulaires pour modifier leurs fonctions sont les mutations génétiques qui affectent l'ADN (comme la modification d'une recette de cuisine) et l'édition de l'ARN (qui agit comme un chef de cuisine). Si le corps humain a abandonné la deuxième technique car désordonnée et sujette à erreur (voir plus bas), en revanche les généticiens l'ont exploitée au point qu'ils peuvent aujourd'hui modifier une seule base (nucléotide) ou lettre du génome. Cette technique utilise l'enzyme appelée CRISPR (lire "crispeur-Cas9" signifiant Clustered Regularly Interspaced Short Palindromic Repeats ou courtes répétitions palindromiques groupées régulièrement espacées) associée à la protéine Cas9, d'où son nom complet de CRISPR-Cas9. C'est une sorte de ciseaux génétiques dont l'usage s'est généralisé car il permet de manipuler l'ADN et créer des OGM et donc potentiellement de remplacer les pathogènes à l'origine de maladies génétiques héréditaires comme la SLA dont fut victime Stephen Hawking ou la démence fronto-temporale (FTD) parmi des centaines d'autres maladies handicapantes ou dégénératives. CRISPR fut découvert en 1987 par l'équipe de Yoshieumi Ishino en étudiant la fameuse bactérie Escherichia coli. Il consiste en une séquence d'ARN de 20 à 40 paires de nucléotides faisant partie du système immunitaire procaryote. Il confère à l'organisme une résistance aux éléments génétiques étrangers tels que ceux présents dans les plasmides (une molécule d'ADN présente dans le cytoplasme) et les phages (des virus n'infectant que les bactéries) en apportant une forme d'immunité acquise. Lorsque l'organisme détecte un ADN étranger, l'ARN hébergeant la séquence aide les protéines Cas associées à CRISPR à reconnaître et à couper l'ADN exogène grâce à leur enzyme (endonucléase). D'autres protéines Cas guidées par l'ARN coupent ensuite l'ARN étranger qui est éliminé. Les CRISPR sont présents dans environ 50% des génomes bactériens séquencés et près de 90% des archées séquencées. Quand on sait qu'aujourd'hui l'identification d'un gène suspect avec CRISPR ne prend que quelques jours contre plusieurs semaines auparavant et même plusieurs mois en 1996, on comprend tout l'intérêt de cet outil. Non seulement, il localise le nucléotide affecté mais grâce à son enzyme Cas il peut aussi couper la base et la remplacer ou la repositionner sur le bon gène. A
voir : CRISPR/CAS9 : une méthode révolutionnaire,
Inserm A lire : Enzymes agissant sur les acides nucléiques, Médecine Sorbonne Université
Parmi les résultats encourageants, en 2017 Emmanuelle Charpentier de l'Institut Max Planck (MPIfIB) et Jennifer Doudna de l'Université de Berkeley ont annoncé dans la revue "Nature" être parvenus à modifier dans un embryon humain in utero des gènes porteurs de la cardiomyopathie hypertrophique, une maladie héréditaire. En 2016, pour la première fois CRISPR fut également utilisé lors d'un test clinique pour traiter une forme agressive de cancer du poumon à l'Hôpital de West China à Chengdu en Chine, et une expérience similaire fut conduite aux Etats-Unis en 2017. Actuellement des équipes de chercheurs testent CRISPR sur des globules blancs (lymphocytes T) pour les aider à combattre les cellules cancéreuses touchant les poumons (cf. Nature, 2018), sur des moustiques OGM afin qu'ils transmettent moins facilement ou plus du tout le paludisme - la maladie fait encore 1 million de victimes par an - (cf. PLOS, 2018), sur des mammifères comme des souris pour tester s'il est possible de tuer des espèces invasives en l'espace d'une ou deux générations (cf. Nature, 2018), dans des tests de dépistage pour détecter le Covid-19 et à terme il servira pour combattre le VIH (cf. NEJM, 2029). Les applications génétiques sont tellement nombreuses dans le domaine médical et dans l'industrie alimentaire que depuis 2018 une nouvelle revue scientifique est dédiée aux travaux de recherche sur l'outil CRISPR, "The CRISPR Journal". Il faut toutefois éviter de jouer à l'apprenti-sorcier comme l'a rappelé le généticien Alexander G. Bassuk de l'Université d'Iowa et ses collègues dans la revue "Nature Methods" en 2017. En effet, la technique CRISPR utilisée in vivo dans les essais cliniques peut induire des centaines de mutations involontaires (jusqu'à 1500 mutations sur un seul nucléotide et plus de 100 suppressions et insertions importantes) non prédites par les algorithmes informatiques. Deux autres études indépendantes publiées dans "Nature Medicine" en 2018 ont également montré que si CRISPR est un outil prometteur dans la thérapie génique, il peut indirectement déclencher de nouveaux cancers, même lorsqu'il est utilisé pour remplacer des gènes cancérigènes (dont p53). Bien entendu, les chercheurs travaillent actuellement à améliorer le système CRISPR pour augmenter l'efficacité de l'édition génomique sans effet de bord. Nous verrons à propos du génie génétique que ces manipulations du génome soulèvent également des questions éthiques notamment concernant l'impact des OGM sur l'environnement et la santé humaine. Nous nous focaliserons néanmoins avant tout sur les OGM du secteur alimentaire, les manipulations génétiques dans le cadre de la santé humaine étant encore balbutiantes et fortement réglementées. L'édition de l'ARN En principe le corps humain réalise rarement l'édition de ses gènes par CRISPR ou autres ciseaux génétiques car comme nous l'avons expliqué, comme la plupart des créatures multicellulaires, pour s'adapter à une nouvelle condition environnementale, notre organisme préfère la mutation génétique, c'est-à-dire la modification de l'ADN (qui assure la fonction de la recette) par le biais de l'ARN (qui assure la fonction de chef cuisinier). C'est une méthode séculaire éprouvée et plus sûre pour appliquer une modification génétique à grande échelle en minimisant les risques grâce à un système élaboré de correction d'erreur qui intervient lors de la transcription.
Mais bien que l'ARN ne soit qu'un exécutant, comme dans le cas de CRISPR parfois il improvise la création de protéines grâce à l'édition de l'ARN permettant à l'organisme de s'adapter sans devoir subir une mutation génétique. Mais ce mécanisme étant archaïque, il présente des inconvénients car il peut engendrer des erreurs génétiques (qui ne changent pas la nature du génome) et son action est désordonnée, provoquant plus de dysfonctionnements qu'elle n'en résout. C'est la raison pour laquelle dame Nature a largement abandonné cette méthode bien que la plupart des organismes disposent toujours des enzymes nécessaires à l'édition de l'ARN. Et c'est heureux pour nous car sans cette "vieille" méthode alternative nous perdrions un bel outil de thérapie génique. A ce jour, seuls quelques organismes marins usent et abusent de l'édition génomique dont les céphalopodes comme l'ont découvert Eli Eisenberg et ses collègues. En 2015, les chercheurs ont expliqué dans la revue "eLIFE" que le calamar a édité plus de 60% de l'ARN de son système nerveux, principalement celui affectant la physiologie de son cerveau, probablement pour s'adapter aux nouvelles conditions de son milieu (température, salinité, pH, polluants, etc). En 2017, dans la revue "Cell", la même équipe de chercheurs rapporta l'observation du même phénomène d'édition génomique chez au moins deux espèces de poulpe et chez la seiche, cette dernière effectuant couramment cette opération. En revanche, le Nautile ne dispose pas de cette faculté d'édition. On savait que la pieuvre peut résoudre des problèmes complexes et est même capable d'utiliser des outils, de dévisser des flacons et d'apprendre en regardant agir ses congénères, et on peut supposer que l'édition de son ARN participe à son intelligence. Le nombre d'acides aminés Pourquoi tous les êtres vivants n'utilisent que 7 à 13 acides aminés sur les 20 qu'ils possèdent pour fabriquer leurs protéines ? Seuls les virus comme le Covid-19 utilisent 20 acides aminés. La question est longtemps restée une énigme jusqu'à ce que le biochimiste évolutionnariste Matthias Granold de l'Université Gutenberg et ses collègues répondent à cette question dans un article publié dans les "PNAS" en 2018.
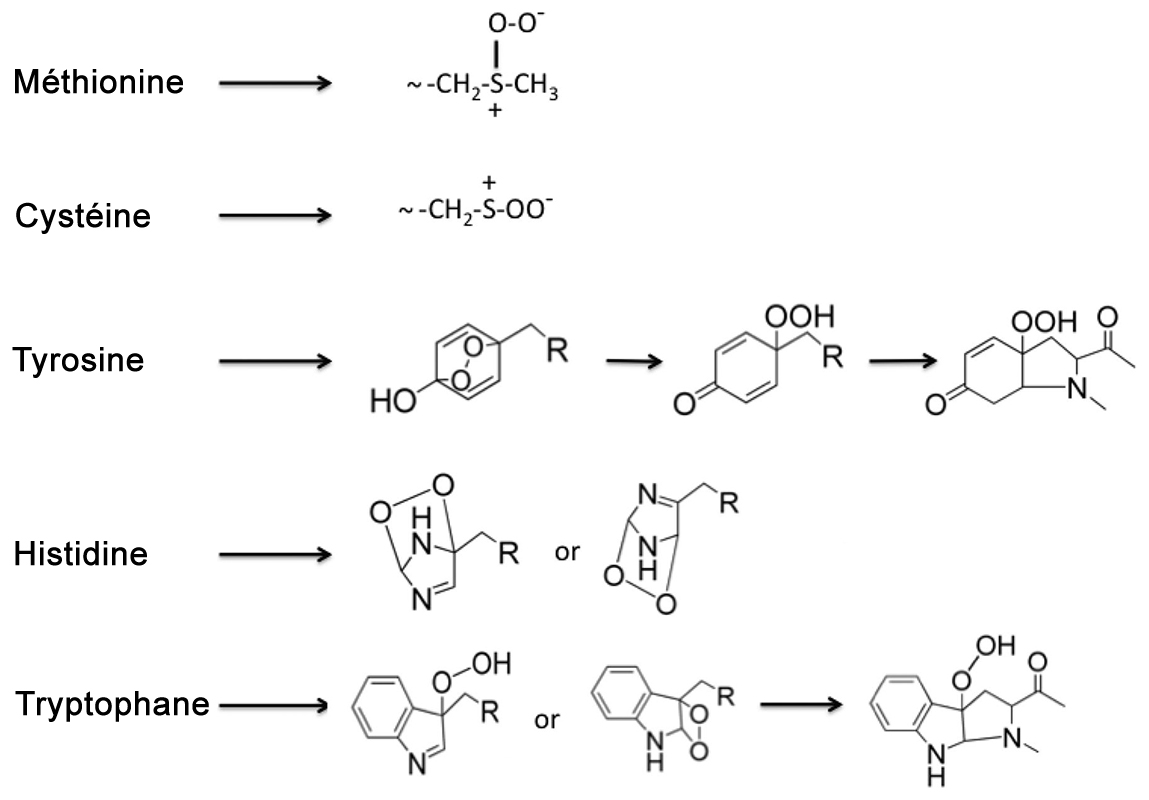
Selon les chercheurs, il faut chercher la réponse dans la composition chimique de l'atmosphère à l'époque des premiers micro-organismes. En effet, il y a plus de 3.8 milliards d'années, grâce à la photosynthèse les premières formes de vie consommaient le gaz carbonique atmosphérique et rejetaient de l'oxygène. Puis, entre 2.45 et 2 milliards d'années, la Terre connut ce qu'on appelle la "Grande Oxydation" durant laquelle le taux d'oxygène augmenta fortement et commença à avoir des effets oxydants (rappelez-vous l'oxydation du fer qui se transforme en rouille). L'oxygène est capable de se lier à d'autres molécules dont il capture plusieurs électrons, créant soit des molécules dans un état oxydé (lié à l'oxygène comme la rouille) soit des radicaux libres (ions) toxiques pour les organismes; dans ce cas l'oxygène devient un poison et à forte concentration il l'est toujours pour les animaux (cf. les effets de l'eau oxygénée). Dans cette atmosphère devenue chimiquement empoisonnée, les organismes n'ont pas eu d'autre choix que de s'adapter ou périr. Pour s'adapter à ce changement environnemental, par sélection naturelle les sept plus récents acides aminés se sont greffés progressivement sur la machinerie génomique des organismes. Le but n'était pas de participer au métabolisme ou à la physiologie cellulaire mais d'empêcher l'action des radicaux libres. Ainsi, les chercheurs ont démontré que certains parmi les nouveaux acides aminés dont la méthionine (M ou MET), le tryptophane (W ou TRP) et la sélénocystéine (U ou SEC) ont pour but de capturer ces molécules toxiques et de les neutraliser sans provoquer de stress ou de dommage dans l'organisme. En utilisant ces nouveaux acides aminés, les organismes ont pu exploiter cet oxygène pour respirer. Autrement dit, la vie a trouvé le moyen de transformer un problème en opportunité pour survivre et se développer; le "poison" oxygène est devenu une denrée vitable mais dont il ne faut pas abuser. Et si nous étendions le code génétique... Inversement, on peut se demander pourquoi tous les organismes utilisent à peine 20 acides aminés pour fabriquer leurs protéines et pourquoi presque toujours les 4 mêmes désoxyribonucléotides A, G, T, C pour fabriquer l'ADN et les 4 mêmes ribonucléotides A, G, U, C pour fabriquer l'ARN ? Ils disposent pourtant d'un assortiment de 64 codons... Cette question pertinente soulève un profond mystère que les biologistes ne parviennent pas à élucider. Hasard, nécessité ou y a-t-il une autre explication ?
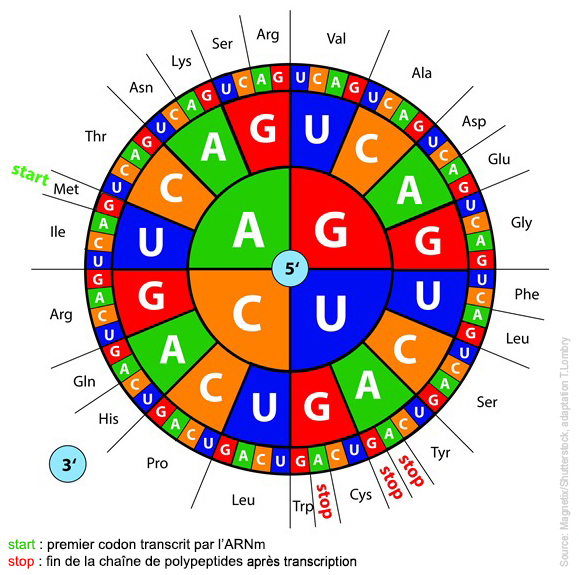
On sait depuis 1977 que certains cyanophages utilisent les bases Z, G, T, C car la liaison hydrogène Z-T est plus stable que la liaison A-T. Elle permet aux virus porteurs du "génome Z" de mieux survivre dans les environnements chauds et difficiles et d'échapper aux défenses immunitaires des bactéries. Pour le reste c'est la grande inconnue. On y reviendra. Nous savons que certains acides aminés inhabituels comme la sélénocystéine et la pyrrolysine, des versions légèrement différentes de la cystéine et de la lysine, sont utilisés par certains organismes. Nous savons également que le monde vivant utilise tous les codons d'une manière ou d'une autre. Certains des 64 codons sont redondants, codant pour le même acide aminé, tandis que trois d'entre eux sont des codons inutiles, dépourvus de sens; ils ne codent pour aucun acide aminé. En revanche, pour le ribosome ils sont les bienvenus car il les utilise comme marqueurs signifiant "stop, fin de la séquence". C'est par exemple le cas du codon stop ambre UAG. Des expériences ont été réalisées en 2004 par l'équipe du professeur Peter G. Schultz du Scripps Research Institute sur des bactéries Escherichia coli en leur ajoutant deux acides aminés non naturels, O-méthyle-L-tyrosine et L-homoglutamine. Comme on s'y attendait, E.coli a utilisé ces acides aminés comme s'ils appartenaient à son patrimoine. Pourquoi dans ce cas dame Nature n'en fait pas autant, comptant sur les mutations aléatoires et la diversité pour améliorer (en principe) ses créations ? Ce processus évolutif qui présente parfois des effets fatals ou inattendus lorsque les gènes s'expriment sans contrôle, sont dédoublés ou triplés (avortement, descendance stérile, déformation, gigantisme, maladies, etc) ferait-il partie d'une stratégie ? Sans évoquer un "moteur divin", si dame Nature préfère l'harmonie et les lois les plus simples, voilà autant de contre-exemples qui peuvent parfois conduire à l'extinction d'une espèce, tout le contraire de son objectif initial, en admettant qu'elle en ait un. Le mystère demeure. Si nous trouvions l'animal à l'origine du dilemme de l'oeuf ou de la poule, nous pourrions peut-être répondre à cette question. Pour y parvenir nous devons essayer de comprendre comment la vie est apparue sur Terre et, si nous découvrons ses traces ailleurs dans l'univers, par quel miracle elle s'y est développée à partir d'unités simples et a priori inertes. C'est le défi que nous propose de résoudre la bioastronomie et sa branche spécialisée qu'est l'exobiologie. Les bases de la génétique étant définies et même extrapolées, voyons comment fonctionne une cellule et quelle est sa structure interne. Ce sera l'objet du prochain chapitre. Prochain chapitre
|
||||||||||||||||||||||||||||||||||||||||||||||