|
|
|
L'origine et l'avenir de l'Homme
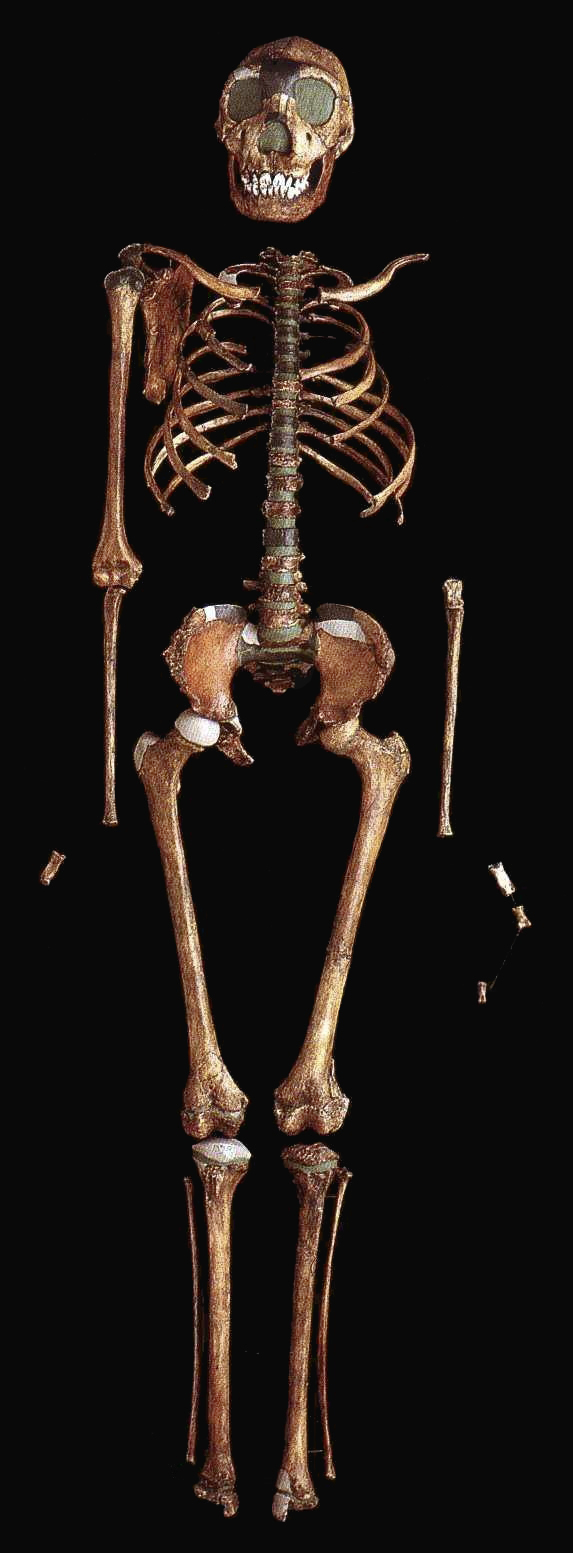
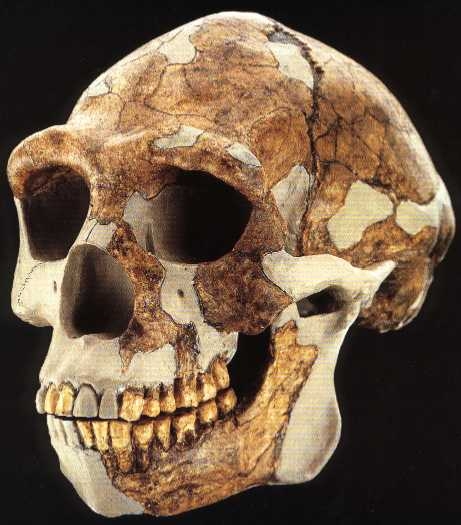
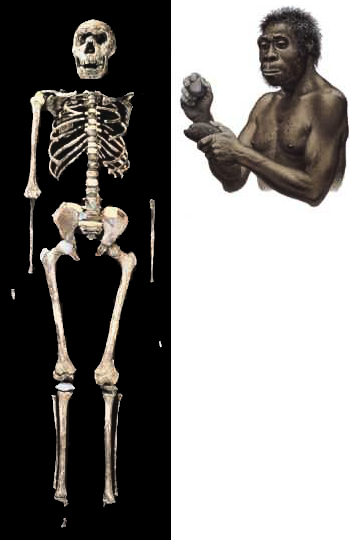
L'Homo erectus : 1.89 - 0.1 million d'années (VIII) Il y a environ 2 millions d'années, une nouvelle espèce d'homininé apparut dans le nord du Kenya : l'Homo erectus ou "l'homme dressé". Il s'agit d'un véritable bipède dont les fossiles remontent entre 1.89 million d'années et 500000 ans voire même 50000 ans pour les spécimens les plus tardifs découverts en Asie du Sud-Est. Cette espèce présente les signes évidents d'une augmentation de la taille de son corps et de son cerveau. D'abord physiquement, le crâne de l'Homo erectus présente deux bourrelets : l’un au-dessus des yeux, formant une longue arcade sourcilière, l’autre au-dessus de la nuque, marquant le point d’attache d’une puissante musculature. Ce n'est pas encore un homme mais à travers les traits bruts de son faciès, quel que soit le lieu géographique où l'époque où il vécut, il présente déjà des signes d'humanité et une intelligence certaine. L'Homo erectus présentait une taille très variable au point qu'il est difficile d'établir une moyenne. La plupart des spécimens mesurent entre 1.45 et 1.70 m et pèsent entre 40 et 70 kg mais il y a beaucoup d'exceptions, autant que dans l'espèce humaine actuelle. La taille de l'Homo erectus est une caractéristique remarquable. Ainsi, l'enfant de Turkana (un Homo ergaster) mesurait 1.50 m à 8 ans et devait mesurer au moins 1.63 m à l'âge adulte (tout dépend de la manière dont les experts établissent leurs mesures). D'autres spécimens mesuraient jusqu'à 1.85 m, ce qui était très grand et inhabituel pour cette époque, les squelettes découverts dans les gorges d'Olduvai en Tanzanie et à Dmanisi, en Géorgie (l'Homo georgicus) pouvant en attester. Il est important de noter que ces variations de taille ne sont pas simplement une augmentation apparue chez les premiers homininés, mais une caractéristique propres à l'Homo erectus, comme elle l'est également chez les humains de notre époque. Parallèlement à l'augmentation de la taille de son corps, le cerveau de l'Homo erectus augmenta sensiblement. Sa capacité crânienne varie entre 750 et 1000 cm3 (plus de 800 cm3 pour l'enfant der Turkana), soit plus de 50% supérieur à celui des Australopithèques (et 60% du volume du cerveau d'un humain moderne). Cette augmentation du volume absolu du cerveau est accompagnée d'une augmentation proportionnelle de l'encéphalisation qui dépassera systématiquement les 1000 cm3 chez les espèces ultérieures, ce qui correspond à la limite inférieure du volume endocrâninen des hommes actuels sans qu'ils soient pour autant marqués par un corps considérablement plus grand que celui des premiers Homo erectus. Le corps plus grand et le cerveau plus volumineux de l'Homo erectus signifient qu'il a besoin de plus d'énergie, et donc de plus de nourriture, que les espèces antérieures. Ses muscles et son cerveau en particulier demandent plus d'énergie pour rester en bon état et performant. L'Homo erectus a donc souvent été représenté comme occupant intensément sa niche écologique au point qu'il a probablement été l'un des premiers hommes à détruire son environnement pour subsister. Mais vu la taille réduite des différentes tribus, son empreinte anthropique resta très faible et temporaire.
Le fait que l'Homo erectus soit assez grand et disposait en particulier de membres assez longs, augmenta son efficacité locomotrice ainsi que le montra l'anthropologue Herman Pontzer de la Cité Universitaire de New York (CUNY) et son équipe dans une étude publiée en 2010 dans le "Journal of Human Evolution". L'Homo erectus pouvait parcourir au sol de bien plus grandes distances en marchant ou en courant que les petits homininés qui l'ont précédé. De plus, son grand cerveau lui offrait de meilleurs facultés pour traiter des informations écologiques complexes à mesure qu'il traversait des territoires à la recherche d'aliments plus nourrissants. L'inventeur de l'industrie de la pierre taillée L'Homo erectus domina la nature non seulement par sa taille mais il était déjà le superprédateur de son temps. Outre la taille de son cerveau, deux autres éléments démontrent qu'il était intelligent et habile. Tout d'abord, il savait fabriquer des outils efficaces. Nous avons des preuves à travers quelques carcasses fossilisées que l'Homo erectus se nourrissait d'animaux de tailles moyenne et grande qu'il chassa ou piéga. Ces carcasses présentent des marques faites par des outils de boucherie. Ce comportement consistant à chasser et décarcasser des animaux constitue un changement écologique comparé à l'attitude des anciens Australopithèques. On constate qu'alors que les plus anciens Homo erectus utilisaient des outils en pierre basiques, typiques de l'industrie lythique de l'Oldowayen (le "Mode 1" tels que des galets aménagés), vers 1.5 million d'années et bien plus encore vers 8500000 ans, les populations d'Homo erectus fabriquèrent des ensembles d'outils plus complexes et typologiquement codifiés qu'on classe aujourd'hui comme le "Mode 2" de l'industrie Acheuléenne, comprenant des bifaces fins et affûtés, des racloirs, des perçoirs, etc. Dans ce contexte on peut affirmer que l'Homo erectus inventa l'industrie de la pierre taillée même si on retrouve ses prémices chez l'Homo habilis. Pour déterminer à quel point ses outils étaient efficaces, des simulations faites par des paléoanthropologues sur de la viande fraîche ont démontré que les racloirs et autres tranchoirs de l'Homo erectus étaient à peine moins performants que les couteaux modernes de nos bouchers. Lors d'une simulation effectuées aux Etats-Unis, on demanda à deux bouchers de dépouiller une carcasse de tous ses morceaux de viande, non pas avec leurs couteaux en acier habituels bien aiguisés mais avec les outils de l'Homo erectus. Ils sont arrivés à bout de leur tâche 4 fois plus lentement que d'habitude. Bien sûr il n'avait pas la méthode de travail ni la dextérité de leurs ancêtres, mais le travail fut accompli, preuve que les outils primitifs en pierre étaient efficaces.
Autre preuve de son intelligence, à la chasse l'Homo erectus savait utiliser la force du groupe pour chasser les animaux et pouvait rivaliser avec les grands fauves qui chassaient en groupe comme les lions, les léopards, les tigres à dents de sabre, les hyènes ou les ours. Encore aujourd'hui, mis à part les tigres à dents de sabre qui ont disparu, les chasseurs considèrent que le léopard (ou le tigre en d'autres endroits) est l'animal le plus féroce de la nature. Non seulement il est agile et rapide mais la taille de ses crocs a de quoi faire reculer le plus téméraire des chasseurs. Ces différents indices témoignent que l'Homo erectus faisait déjà preuve d'une grande intelligence, non obstant le fait qu'il était costaux et courageux. Mais ses aptitudes ne l'ont pas sauvé de l'extinction. A propos des dents justement, le changement écologique associé à l'Homo erectus coïncide avec un changement de la dentition de cette espèce. Comparé aux anciens Australopithèques telles que Abel ou Lucy et aux variétés robustes qui vécurent à son époque (Paranthropus robustus), la dimension de la dentition post-canine (prémolaires et molaires) et la molarisation des prémolaires se sont sensiblement réduites chez l'Homo erectus. Le corps de la mandibule (la partie édentée qui est reliée au crâne) montre aussi une plus grande gracilité ou minceur avec une diminution de la largeur de la structure de mastication.
Selon Ungar et son équipe, l'analyse de ses dents indique qu'à travers les âges et les régions, l'Homo erectus a dû faire face à diverses formes de régimes alimentaires. En outre, la nourriture qu'il mangeait a modifié la chimie de ses tissus corporels (selon l'adage vous êtes ce que vous mangez), y compris la dentine et l'émail de ses dents. Leur analyse radioisotopique a permis de montrer que l'Homo erectus était moins sensible et plus flexible par rapport à la nourriture que ses ancêtres. Enfin, quelle que soit le type d'aliment carné ou végétal dont se nourrissait l'Homo erectus selon les régions géographiques, l'anatomie de ses dents et de ses mâchoires révèlent que son régime alimentaire ne nécessitait pas les mêmes adaptations masticatoires robustes que celles observées chez les homininés plus anciens. Cela laisse supposer que l'Homo erectus était omnivore dès son apparition il y a environ 2 millions d'années et que cette caractéristique a dû émerger un ou deux degrés de parenté avant lui, probablement chez des Australopithèques tardifs. Concernant le dimorphisme sexuel, les différences physiques entre les mâles et les femelles sont difficiles à établir du fait de la grande variabilité de la taille des Homo erectus, au moins équivalente à celle qu'on observe aujourd'hui dans la population humaine. On ne peut donc pas affirmer que les mâles étaient systématiquement plus grands que les femelles ou d'un dimorphisme différent de celui qu'on observe aujourd'hui entre l'homme et la femme. Toutefois, si l'Homo erectus présentait un dimorphisme sexuel supérieur à celui de l'Homo sapiens, cela aurait eu pour conséquence que la compétition des mâles pour les femelles dépendait plus de la taille du corps qu'il ne l'est aujourd'hui. Quant à l'acquisition du langage, sur lequel nous nous attarderons plus loin, il est difficile de l'évaluer directement à partir des fossiles. Des tentatives d'analyses ont été faites sur différents aspects du système nerveux central, y compris sur la dimension du canal cervical (une approximation de la taille de la moelle épinière) et des détails endocrâniens, mais aucune découverte significative n'apporte la preuve que l'Homo erectus était capable d'un quelconque proto-langage. En revanche, des analyses génétiques d'échantillons prélevés chez l'homme de Néandertal et chez l'Homo desinova ont montré que le gène FOX2 a joué un rôle dans le développement du langage. Ce gène serait apparu au moins dès le Pléistocène Moyen, et donc il est tout à fait possible que l'Homo erectus en ait hérité par sa lignée ancestrale humaine. Malheureusement, les chercheurs ne disposent d'aucun ADN d'Homo erectus pour tester cette hypothèse. Cela ne signifie pas que l'Homo erectus était génétiquement capable de parler, mais comme le prouvent les études anatomiques, nous n'avons aucune preuve non plus permettant de rejeter cette idée. Pourquoi l'Homo erectus s'est-il éteint ? Parmi les questions en suspens, on se demande pour quelle raison l'Homo erectus a-t-il disparu ? D'abord, il faut rappeler que cette espèce vécut à une époque où le climat était en pleine évolution et qu'il n'est jamais facile de survivre quand on est adapté à un environnement verdoyant et que soudainement (en quelques milliers d'années) il se transforme en steppe puis en désert. Mais ce n'est pas le seul facteur qui contribua à son extinction. Dans une étude publiée en 2018 dans la revue "PLoS One", Ceri Shipton de l'Université Nationale Australienne (ANU) et ses collègues ont montré que l’Homo erectus disparut (du moins en Arabie) en partie du fait qu'il était "paresseux". En effet, des fouilles archéologiques entreprises en 2014 sur le site de Saffaqah proche de Dawadmi situé au centre de l'Arabie Saoudite ont révélé que l'Homo erectus utilisait des "stratégies de moindre effort" pour fabriquer ses outils et collecter ses ressources et cela affecta sa survie. Cette "paresse" associée à une incapacité à s'adapter à un climat en évolution a probablement joué un rôle dans la disparition des Homo erectus. Selon Ceri Shipton, "pour fabriquer leurs outils en pierre, ils utilisèrent toutes les roches qu’ils pourraient trouver autour de leur camp, qui étaient de qualité relativement médiocre par rapport à celles utilisées ordinairement par les hommes du néolithique. Sur le site, nous avons découvert un grand éperon rocheux de pierre de qualité situé à une courte distance de leur camp mais qu'ils n'avaient pas exploité. Plutôt que de monter sur la colline, ils n'utilisèrent que les éclats qui étaient tombés et se trouvaient au sol. Quand nous avons examiné l’affleurement rocheux, il n’y avait aucun signe d’activité, d'artefacts et d'extraction de la pierre. Ils savaient que les pierres étaient là, mais du fait qu'ils disposaient de suffisamment de ressources, ils semblent s'en être désintéressés". Cette attitude contraste avec celle des hommes des industries lithiques des périodes ultérieures, notamment les premiers Homo sapiens et Néandertaliens, qui escaladaient des montagnes pour trouver des pierres de bonne qualité et les transportaient sur de longues distances.
Les chercheurs estiment que l’incapacité de progresser sur le plan technologique à mesure que leur environnement s'est asséché et devint désertique, a également contribué à la disparition des Homo erectus. Selon Shipton, "non seulement ils étaient paresseux, mais ils étaient aussi très conservateurs. Les échantillons de sédiments ont montré que bien que leur environnement changeait, ils ont continué à faire exactement la même chose avec leurs outils. Il n'y a pas eu de progression du tout et ils ont toujours trouvé leurs outils près des lits de rivières aujourd'hui à sec. En fin de compte l'environnement est devenu trop sec pour eux." Maintenant la question est de savoir si cette conclusion applicable à une petite population d'Homo erectus d'Arabie centrale peut être généralisée comme le sous-entend Shipton aux millions d'Homo erectus vivant à travers la planète ? Il paraît présomptueux et prématuré de l'affirmer. Extensions géographique et temporelle On prétend que l'Homo erectus est la plus ancienne espèce du genre humain à présenter autant de qualités similaires aux nôtres (bipédie, grand cerveau, dextérité, etc), bien que ses fossiles présentent un certain nombre de traits spécifiques et distincts de ceux des êtres humains actuels. C'est aussi l'espèce qui suivit le plus long parcours évolutif pour aboutir à ce que nous sommes ou presque (nous verrons que l'Homo sapiens ne descend pas directement de l'Homo erectus).
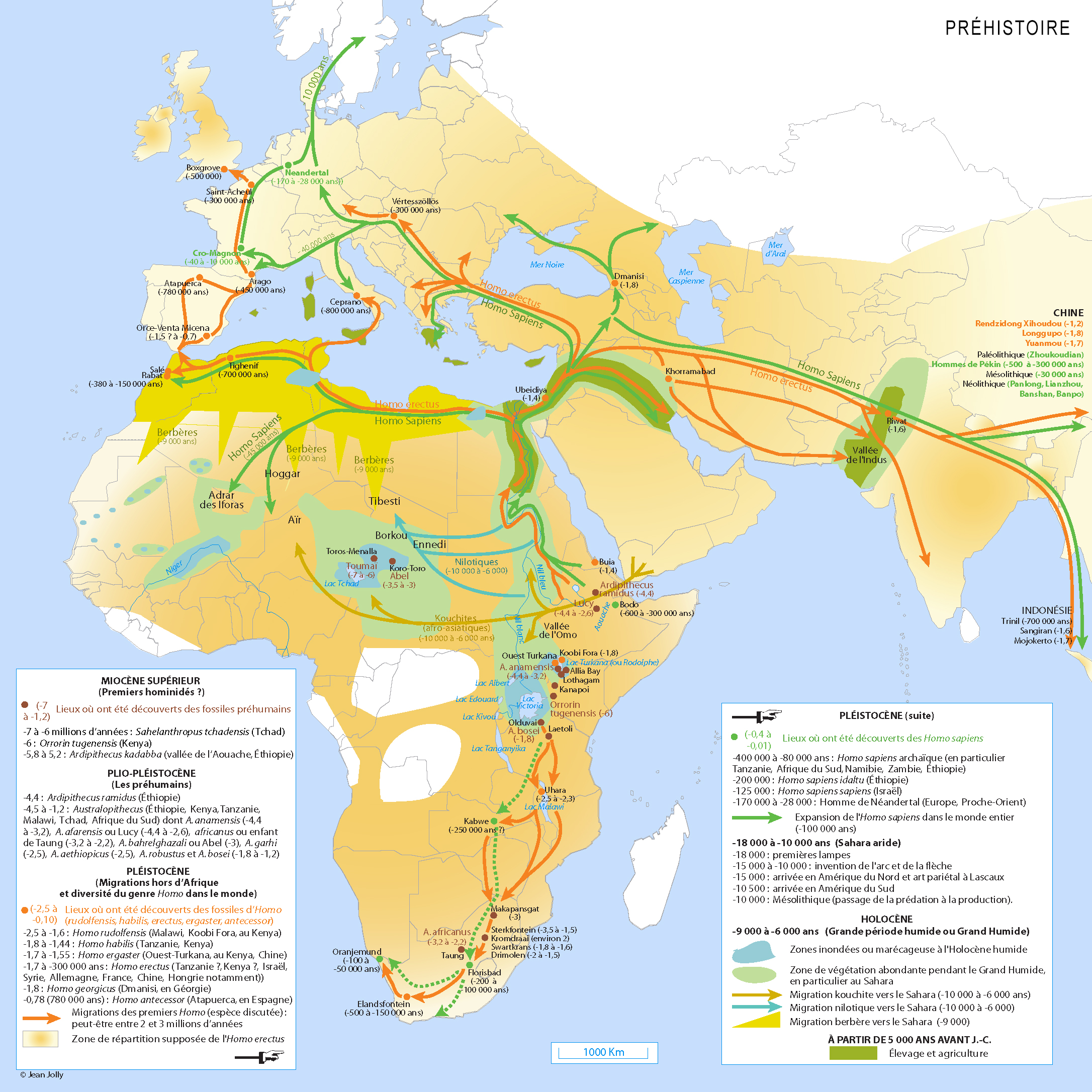
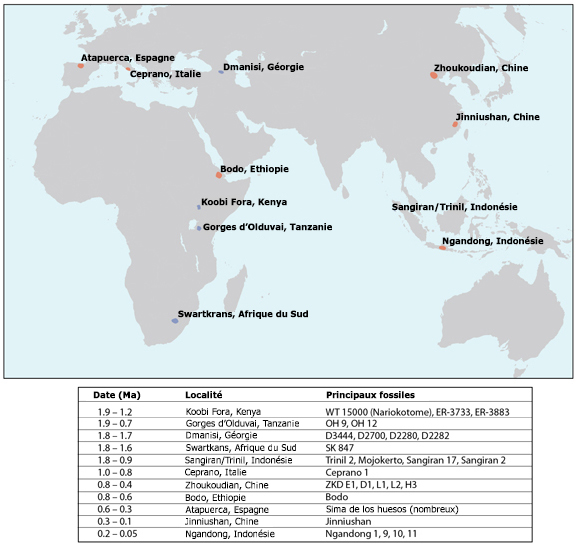
Si l'Homo erectus présente une remarquable adaptation au climat, il est aussi souvent considéré comme le premier homininé d'une lignée cosmopolite, c'est-à-dire la première espèce humaine dont la distribution géographique s'est étendue au-delà d'une seule région continentale. Comme les fossiles d'homininés plus anciens, on trouve des fossiles d'Homo erectus en divers lieux d'Afrique parfois distants de plusieurs milliers de kilomètres, mais également dans de nombreux sites distribués à travers toute l'Eurasie comme le montre les cartes présentées ci-dessous. Non seulement l'espèce connut une distribution géographique quasi mondiale, mais elle s'est également étendue dans le temps sur pratiquement 1.9 million d'années, ce qui en fait l'espèce humaine ayant eu la plus longue longévité. Apparu pour la première fois dans le bassin du lac Turkana, au Kenya (considérés aujourd'hui comme des Homo ergaster, voir plus bas), dans sa course à travers monts et vallées, juste armé de bifaces et de lances, l'Homo erectus fut le premier à parcourir l'Afrique du Sud et de l’Est. Entre 1 million d’années et 700000 ans il remonta vers les zones tropicales, traversant l’Afrique et l'Eurasie pour conquérir de nouveaux territoires de chasses. On retrouve des sous-espèces de l'Homo erectus des Tropiques jusqu'au Grand Nord, de l'Espagne en passant par l'Indonésie jusqu'en Chine entre 750000 et 300000 ans. Les dernières populations d'Homo erectus ont survécu pratiquement jusqu'à la fin du Pléistocène dans le sud-est de l'Asie, les derniers fossiles ayant été trouvés à Ngandong, en Indonésie, et remontent entre 100000 et 50000 ans. On ne sera donc pas étonné d'apprendre que la population d'Homo erectus comptait déjà plusieurs millions d'individus. L'Homo erectus est une espèce intéressante à plus d'un titre qui met au défi les paléoanthropologues essayant d'interpréter la variété des fossiles découverts dans le contexte d'une répartition géographique et temporelle si étendue. Nous allons y revenir. L'expansion de l'Homo erectus à travers une grande variété d'environnements suggère également qu'il y eut une modification d'ordre écologique dans la lignée des premiers homininés, un changement qui a certainement marqué son empreinte dans l'évolution des espèces et modelé la variabilité qu'on observe dans toute la lignée humaine depuis plusieurs millions d'années. Concernant la question de la répartition géographique de l'espèce, certains chercheurs souhaitent que la désignation Homo erectus soit limitée aux seuls fossiles découverts en Asie de l'Est et du Sud-Est, conformément à l'origine des premiers fossiles du taxon. Dans ce cas, la majorité de ses représentants vécurent de la fin du Pléistocène Inférieur jusqu'au Pléistocène Moyen (~1.4-0.2 million d'années). Les fossiles plus anciens découverts dans la partie ouest de l'Asie (à Dmanisi en Georgie) et en Afrique (à Koobi Fora au Kenya) sont similaires à l'Homo erectus classique d'Asie mais présentent des traits ancestraux, qu'on pourrait considérer comme une lignée séparée : l'Homo ergaster. Certains chercheurs ont également scindé l'espèce en fonction de régions plus réduites et assigné par exemple à la population d'Asie de l'Ouest une espèce à part entière : l'Homo georgicus. Mais c'est loin d'être prouvé et de faire l'unanimité (voir plus bas). De même, les fossiles datant du Pléistocène Moyen découverts en Europe ont été attribués à une troisième espèce : l'Homo heidelbergensis. Classée ainsi, la niche écologique occupée par chaque espèce est plus limitée, conduisant à l'isolement et finalement à la spéciation des différentes populations régionales.
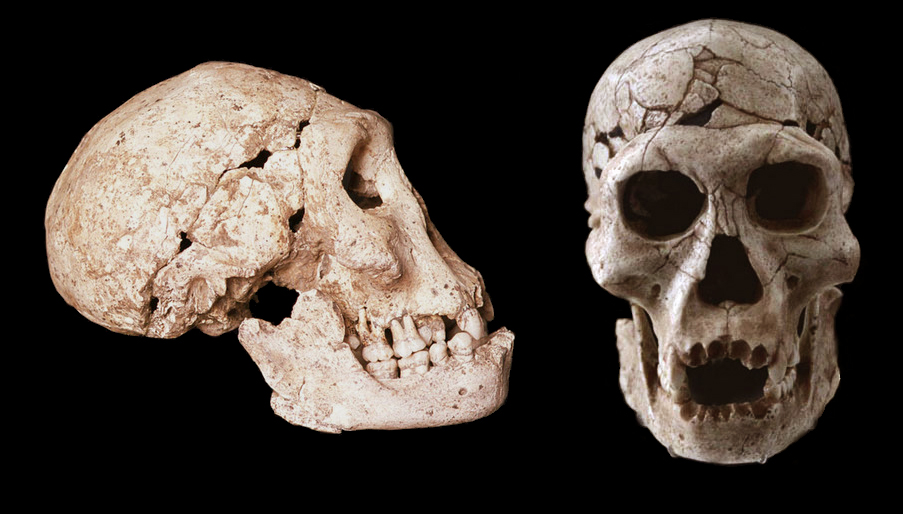
La plus grande partie de la variabilité qu'on observe aujourd'hui dans la population humaine est le résultat de changements génétiques relativement récents survenus au cours des 100000 dernières années de notre histoire. En revanche, les schémas de variabilité de l'Homo erectus se sont déroulés sur une échelle de temps supérieure à 1 million d'années et furent probablement différents de ceux qu'on observe aujourd'hui. Tout le défi des chercheurs est d'expliquer cette variabilité chez l'Homo erectus tout en essayant de comprendre comme les forces de l'évolution ont opéré à travers de telles échelles de temps. Bien sûr beaucoup de questions restent ouvertes qui nécessitent une approche multidisciplinaire et dont on espère avoir quelques éléments de réponses dans les années à venir. En résumé, l'Homo erectus représente une transformation significative des premiers homininés que sont les Australopithèques vers une espèce beaucoup plus proche des hommes modernes. Par rapport à son ancêtre, l'Homo erectus est plus grand, plus intelligent et plus adapté pour survivre dans les différents habitats que présente alors le monde en plein changement climatique. Malgré le gouffre temporel qui nous sépare de l'Homo erectus, bien qu'il était probablement incapable de parler mais tout à fait capable de gesticuler, ce sont des caractéristiques dans lesquelles nous nous reconnaissons. Disposant de capacités cognitives évoluées, écologiquement actif et ayant un comportant hautement adaptatif (mais parfois paresseux), l'Homo erectus a marqué un changement majeur dans l'évolution de l'humanité qui eut des effets du Pléistocène à aujourd'hui. Par de nombreux côtés, notre espèce n'est qu'une version mise à jour de notre ancêtre Homo erectus qui a compris qu'elle devait tout miser sur son intelligence pour survivre et s'adapter à son environnement. A lire : Homo erectus made world's oldest doodle 500,000 years ago, Nature, Déc 2014 Les plus vieux griffonnages (sur coquillage) Les espèces controversées Au cours de leurs recherches, les paléoanthropologues ont découvert de nombreux squelettes ressemblant à ceux de l'Homo erectus mais généralement plus primitifs par certains détails qui ont conduit leur découvreur à les classer dans une nouvelle espèce d'homininé. Toutefois, leur hypothèse n'est pas soutenue par tous les chercheurs qui n'y voient généralement que la grande variabilité des Homo erectus. Parmi ces nouvelles espèces contemporaines de l'Homo erectus mais qui font débat, citons : - 1. l'Homo ergaster vivant en Afrique - 2. KNM-ER 42700 et 42703 - 3. l'Homo georgicus vivant en Géorgie - 4. le Sinanthropus pekinensis ou "l'homme de Pékin" vivant en Chine. Disons tout de suite à propos de "l'homme de Pékin" (cf. le site de l'UNESCO) que la majorité des scientifiques a finalement préféré le considérer comme une sous-espèce de l'Homo erectus : l'Homo erectus pekinensis. Après la description des deux espèces suivantes, on pourrait donc aussi les considérer comme deux sous-espèces de l'Homo erectus, une théorie que paradoxalement les découvreurs eux-mêmes supportent. Mais cela reste surtout un débat taxonomique qui n'intéresse que les experts. 1. L'Homo ergaster : 1.8 - 1 million d'années C'est en 1991 que Bernard Wood, alors à l'Université de Liverpool proposa de distinguer les Homo erectus découverts en Eurasie de l'espèce vivant exclusivement en Afrique qu'il baptisa Homo ergaster. Selon Wood, "cette espèce présente des traits proches de ceux des Homo erectus mais plus généralistes et plus primitifs que les groupes indonésiens et chinois." Cette controverse débuta en 1984, lorsque Kamoya Kimeu, responsable d'une équipe de paléoanthropologues dirigée par Richard Leakey et surnommée le "Gang des Hominidés" ("Hominid Gang") découvrit à Nariokotome près du lac Turkana au Kenya (près des sites de KNM-ER 1470 et KNM-ER 3733), un petit squelette qui sera dénommé prosaïquement KNM-WT 15000. L'ouverture étroite de son pelvis et sa dentition permirent de déterminer qu'il s'agissait d'un individu mâle et juvénile. Le spécimen fut surnommé le "garçon de Nariokotome" ou "garçon de Turkana".
Lors de la première analyse, l'âge de l'enfant fut estimé entre 7.5 ans et 15 ans. Un réexamen fut réalisé par Ronda.R. Graves et ses collègues sur base d'un modèle de croissance propre aux premiers humains (qui est plus court que chez l'homme moderne). Leur analyse publiée en novembre 2010 dans le "Journal of Human Evolution" indique que l'enfant serait âgé de 8 ans et devait mesurer 1.63 m à l'âge adulte. Le squelette remonte au Pléistocène inférieur, il y a environ 1.6 million d'années. C'est actuellement le squelette humain le plus complet remontant à cette période. En 1995, les squelettes fossilisés de deux homininés de la même espèce comprenant leur crâne et leurs os longs furent découverts dans le désert de Danakil en Érythrée. L'un des spécimens fut surnommé "l'homme de Buya". Il vécut il y a 1 million d'années. Jusqu'à preuve du contraire, "l'enfant de Turkana" et "l'homme de Buya" appartiennent à l'espèce Homo ergaster. Elle vécut dans l'est de l'Afrique durant une période qui s'étendit entre 1.8 et 1 million d'années. Elle cohabita probablement avec l'Homo habilis qui vivait également dans la région du lac Turkana. Selon les défenseurs de ce taxon, l'Homo ergaster serait l’ancêtre de l'Homo erectus dont il occupa la niche en Afrique de l'Est et probablement de l'Homo antecessor qui migra vers l'Europe sur lequel nous reviendrons.
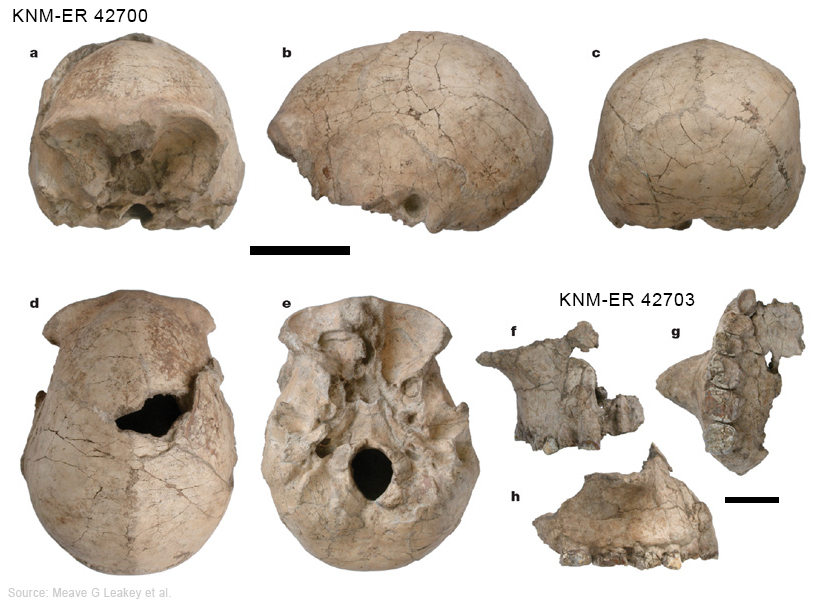
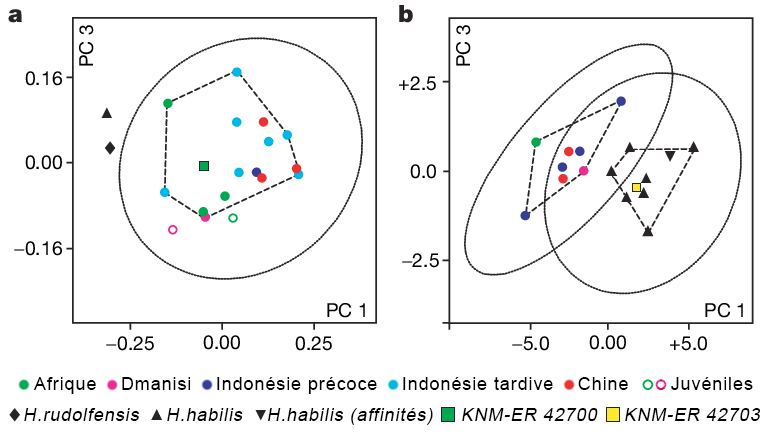
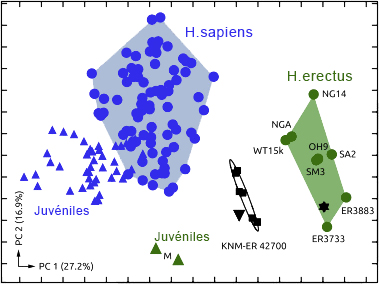
L'Homo ergaster mesurait entre 1.55 et 1.70 m et pesait entre 50 et 65 kg. Sa capacité endocrânienne varie entre 850 et 1000 cm3, un volume important qui demande assez bien d'énergie, obligeant vraisemblablement les individus à chasser et consommer de la viande. Anatomiquement, l'empreinte endocrânienne présente une forte irrigation sanguine et une asymétrie semblable à celle d'un cerveau moderne. Les os pariétaux sont presque parallèles. L'Homo ergaster présente un faciès très archaïque, un nez plat et une mâchoire massive et très prognathe. Le dimorphisme sexuel est en revanche plus réduit que chez l'Homo habilis. Ses membres inférieurs plus longs que ceux d’Homo habilis, ses jambes très peu arquées et ses pieds présentant des orteils parallèles en font un bon marcheur capable de courir. Les hanches des femelles sont en revanche plus larges que cellles de l’Homo sapiens femelle. L'Homo ergaster vivait en forêt, non loin des fleuves et parmi une faune très diversifiée. L'analyse stratigraphique identifia 21 espèces de vertébrés et 16 espèces de mammifères. Comme l'Homo habilis, l'Homo ergaster utilisa des outils en pierres taillées et notamment des bifaces caractéristiques de l'Acheuléen dont il serait l'inventeur. On a retrouvé de nombreuses traces laissées par ses outils lithiques sur des os d'antilopes, d'hippopotames et même de crocodiles. Les analyses indiquent que l'Homo ergaster parcourait de grandes distances pour sélectionner ses outils et était capable de les tailler avec habileté ce qui impose un savoir-faire. Cela implique une certaine éducation et un apprentissage, et donc le développement des prémices d'une culture héritée des Anciens et transmise aux jeunes. Parmi les chercheurs opposés à cette nouvelle distinction taxonomique, il y a le paléoanthropologue géorgien David Lordkipanidze pour qui "le garçon de Turkana" est en fait "le squelette d'Homo erectus le mieux préservé". 2. KNM-ER 42700 et 42703 Par la suite d'autres squelettes ont complété les collections dont le sommet du crâne ou calvaria de KNM-ER 42700 découvert au Kenya daté de 1.55 million d'années et lune partie du maxillaire de KNM-ER 4703 daté de 1.44 million d'années. Il s'agit de deux spécimens qui se situent respectivement entre l'Homo erectus africain et les premiers hommes d'Asie et entre l'Homo habilis et l'Homo erectus mais dans les deux cas, l'espèce reste inconnue. Fred Spoor et Meave Leakey ne sont pas parvenus à distinguer leur appartenance : "Le crâne de KNM-ER 42700 correspond à 95% à celui de l'Homo erectus mais il est différent de celui de l'Homo habilis et de l'Homo rudolfensis. Le maxillaire de KNM-ER 42703 correspond à 95% à celui de l'Homo habilis mais il est différent de celui de l'Homo erectus." Spoor poursuit : "En examinant les plus petits détails [du fossile KNM-ER 42700], j'ai été obligé de conclure qu'il n'y a pas de séparation claire entre les deux [espèces]. Il présente en effet des caractères typiquement "asiatiques" : une carène sagittale sur l'os frontal et l'os pariétal, des arrangements de la base crânienne […] qui sont reliés avec l'orientation du canal auditif identiques à ceux que Franz Weidenreich avait décrit dans les années 1940 pour l'homme de Pékin." Lee Berger et les paléoanthropologues de l'équipe de Meave Leakey se demandent aujourd'hui si KNM-ER 42700 ne serait pas un Homo naledi vivant au Kenya ? La question est ouverte. A moins que ces deux spécimens apportent les preuves qu'il existait encore d'autres espèces humaines en Afrique... ce qui en soi est tout à fait possible.
Actuellement le débat est toujours ouvert et certains musées nationaux ont choisi délibérément de ne pas mentionner l'Homo erectus dans leurs collections, notamment le célèbre Musée National d'Histoire Naturel du Smithsonian (NMNH) alors que le Musée d'Histoire Naturelle de Londres (NHM) par exemple le mentionne. Ceci dit, l'analyse paléogénétique - à condition de trouver de l'ADN dans ces squelettes - pourra sans doute un jour trancher la question. 3. L'Homo georgicus : 1.85 - 1.78 million d'années Le village de Dmanisi situé à 90 km au sud-ouest de Tbilissi, la capitale de la Géorgie, est connu depuis 1936 pour abriter un important site archéologique depuis que les chercheurs y ont découvert les vestiges de plusieurs occupations humaines remontant entre l'Âge du Bronze et la période médiévale. Ce village se trouve égaement sur la route de la Soie. Les géologues découvrirent que juste avant la première occupation de Dmanisi, la vallée de Mashavera fut envahie par une coulée de lave de 80 à 100 mètres d'épaisseur qui forma le basalte de Mashavera. Ce basalte est à l'origine de la formation d'un lac d'environ 1 km de long situé au sud du site. L'analyse paléomagnétique révéla que la coulée était âgée de 1.8 million d'années. Le site paléontologique est situé à environ 4 km à l'est du village, sur un promontoire s'élevant à environ 80 mètres au dessus de la confluence des rivières Mashavera et Pinezaouri, comme on le voit ci-dessous. C'est à cet endroit qu'en 1983 les chercheurs découvrirent au fond d'anciens puits de stockage de grains les ossements d’animaux dont une dent de rhinocéros (Dicerorhinus etruscus etruscus) remontant au Pléistocène inférieur (2.5-0.8 million d'années). En 1984, au même endroit, le paléoanthropologue Abesalom Vekua et ses collègues découvrirent des pierres taillées prouvant que des homininés y vécurent également. Des fossiles typiques de la faune Villafranchienne indiquaient que la strate remontait entre 1.8-1.7 million d'années.
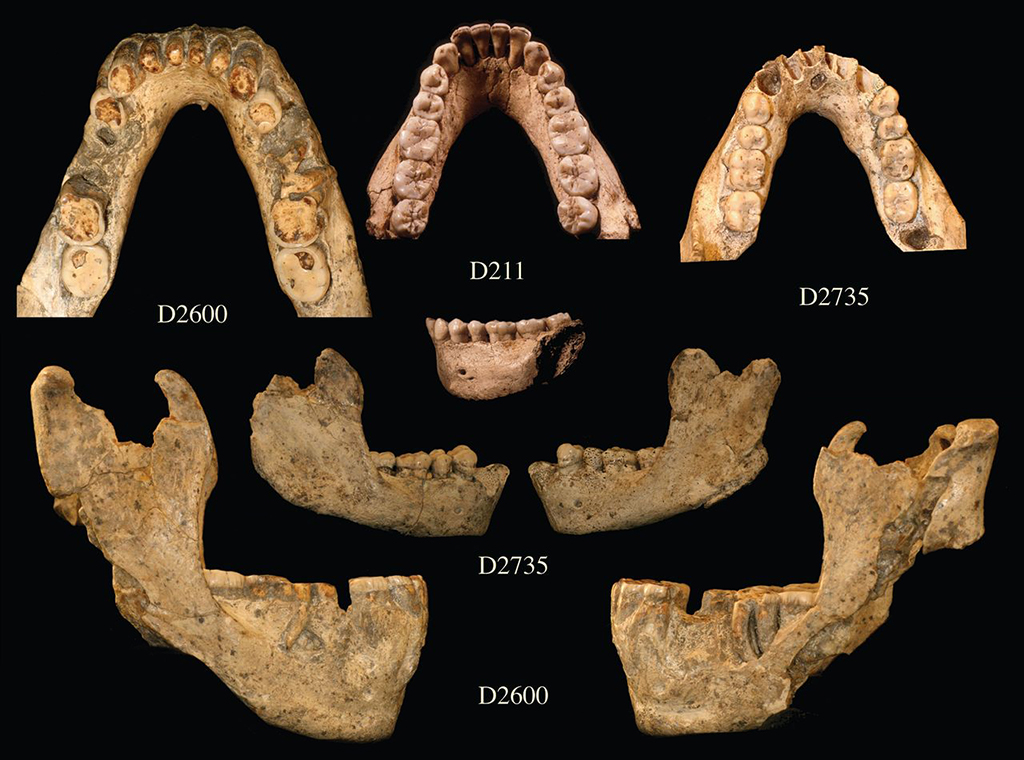
La découverte des outils en pierre était la preuve que des humains avaient déjà quitté l'Afrique à l'époque de l'Homo erectus et avaient déjà peuplé les latitudes Nord. Il fallait à présent trouver des squelettes et identifier à quelle espèce ils appartenaient. Différents centres de recherches (géorgien, allemand, français, américain, espagnol, etc) participèrent à ces études et notamment la Fondation Leakey qui offrit 10% du budget alloué aux chercheurs géorgiens. A partir de 1991, le paléoanthropologue géorgien David Lordkipanidze, directeur du Musée National de Géorgie (GNM) et plusieurs équipes de chercheurs internationaux travaillèrent sur le site et découvrirent dans des tanières les premiers ossements humains constitués d'une petite mandibule (D211) appartenant à un jeune adulte. En 1999, les chercheurs découvrirent les fragments de deux crânes humains (D2282, le crâne d'une jeune femme entre 25 et 30 ans et D2280, la calotte crânienne d'un jeune homme, puis une grande mandibule (D2600) en 2000 présentée ci-dessous. La découverte fut annoncée le 12 mai 2000 dans la revue "Science". Les auteurs estimèrent que les squelettes appartenaient à des Homo ergaster. Poursuivant leurs travaux d'excavation, en 2001 les chercheurs découvrirent un troisième crâne presque complet (D2700) puis sa mandibule (D2735) à 4 mètres de distance et des ossements de trois adultes comprenant notamment des os longs, des côtes, des vertèbres et quelques phalanges. Entre 2003 et 2004, ils découvrirent un tibia et un talus, puis un quatrième crâne édenté en 2005 (D3444) et une mandibule (D3900). Enfin, un cinquième crâne (D4500) fut découvert en 2005. Par chance, la mandibule D2600 s'avéra lui appartenir.
La datation révéla que les fossiles sont âgés entre 1.85-1.78 million d'années. En raison de son intérêt culturel, en 2007 le site de Dmanisi fut inscrit au Patrimoine Mondial de l'Humanité par l'UNESCO. Les résultats des analyses complémentaires furent notamment publiés dans les revues "Nature" en 2007 et "Science" en 2013. Grâce à ces ossements, les chercheurs purent estimer avec précision la taille et la proportion du corps ainsi que la démarche de ces homininés. Le fait que les squelettes aient été trouvés dans des tanières suggère que les individus furent dévorés par de grands prédateurs, une hypothèse qui reste à confirmer. Sur le plan morphologique, selon David Lordkipanidze, les hommes adultes devaient mesurer entre 1.40-1.50 m et peser entre 40-50 kg, soit significativement inférieur aux mensurations moyennes de l'Homo erectus et de l'Homo ergaster qui vécurent à la même époque et avec lesquels ils partagent plus d'un trait en commun.
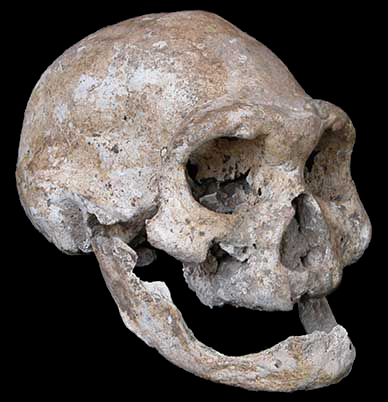
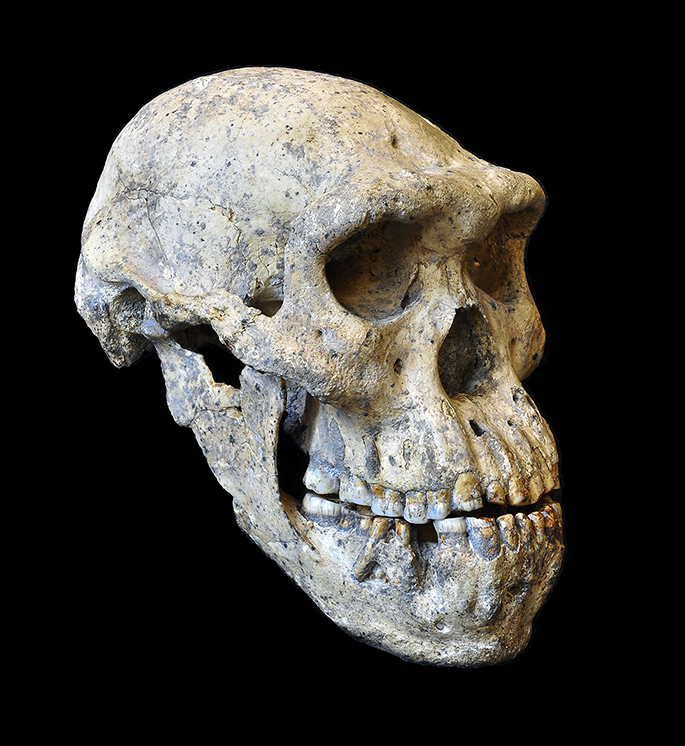
L'analyse du crâne édenté D3444 et notamment de sa maxillaire (mâchoire supérieure) et de sa mandibule (machoire inférieure) suggère que l'individu survécut plusieurs années sans pouvoir mâcher. Cela suppose que les autres membres de cette tribu éprouvaient plus que de la sympathie pour cette personne; ils l'ont nourrie et présentaient donc de la compassion pour elle. Comme le fit l'Homo erectus, ce vieil homme faisait déjà partie d'une communauté soudée où tous les membres partageaient les aléas de la vie, les petits bonheurs comme les malheurs qui pouvaient s'abattre sur l'un ou l'autre individu. Cela confirme que le sentiment d'humanité (sans pour autant parler d'amour du prochain) est apparu très tôt dans l'histoire des hommes, bien avant l'apparition de nos ancêtres directs dont on crut longtemps qu'ils nous avaient légué ce sentiment. Concernant les autres crânes, celui du spécimen D2280 présente un volume endocrânien de 780 cm3, typique de l'Homo erectus ou de l'Homo ergaster. Le volume endocrânien du crâne D2700 présenté ci-dessous à gauche et au centre est d'environ 600 cm3. Sa forme ainsi que sa face courte et plate sont typiques des humains primitifs. Ses canines sont également très apparentes et comme le crâne des autres individus, celui-ci est fuyant, ce qui le rapproche plus des espèces primitives que de l'homme moderne. Le volume endocrânien du crâne D4500 présenté ci-dessous à droite est de 546 cm3. A ce jour, c'est le crâne d'homininé le plus petit et le plus primitif de la période du Pléistocène (en excluant l'Homo floresiensis). Sa taille et sa morphologie font penser qu'il s'agit d'un Australopithèque. De plus, comme celui de D2700, la forme, la face et l’orientation du crâne indiquent qu'il s'agit d'un individu humain primitif. En revanche, la mandibule D2600 a toutes les caractéristiques et notamment les petites canines et le prognathisme du genre Homo. Mais c'est leur variabilité individuelle qui est étonnante; elle est égale ou supérieure aux variations qu'on rencontre chez l'Homo habilis, l'Homo erectus et chez l'Homo rudolfensis. Cette variabilité morphologique incita les chercheurs à examiner les variabilités chez l'homme moderne et les chimpanzés. Ils découvrirent que si les crânes de Dmanisi varient considérablement d'un individu à l'autre, ces diférences ne sont pas plus grandes que celles qu'on observe entre les hommes modernes et entre les chimpanzés.
Les chercheurs ont donc préféré ne pas classer prématurément les spécimens de Dmanisi dans une nouvelle espèce. En effet, en raison de cette forte variabilité entre les spécimens, les squelettes peuvent parfaitement appartenir à des Homo erectus ou à des Homo ergaster parmi d'autres. En revanche, d'autres chercheurs ont relevé des caractéristiques spécifiques majeures comme la mandibule D2600 qui leur suffit pour créer une nouvelle espèce : l'Homo georgicus. Dans ce cas, il serait le descendant de l'Homo habilis et l'ancêtre de l'Homo erectus d'Asie. Mais le chercheur français Antoine Balzeau du CNRS qui participa également à ces analyses, considère que sur base du peu de fossiles existants et donc d'analyses comparées complètes, il est très difficile pour ne pas dire impossible de préciser si ces individus sont des Homo erectus ou d'une autre espèce. En fait, si on va jusqu'au bout du raisonnement, ces variabilités suggèrent que les fossiles assignés jusqu'ici à différentes espèces sur base de leurs spécificités, telle que l'Homo rudolfensis, l'Homo gautengensis, l'Homo ergaster et potentiellement certains fossiles assignés à l'Homo habilis, peuvent être interprétés comme appartenant à la même lignée, celle de l'Homo erectus. C'est finalement David Lordkipanidze qui trancha la question en effaçant pour ainsi dire toutes les variabilités pour ne garder qu'un seul taxon : "Etant donné que jusqu'ici chaque lignée Homo pouvait être définie par des caractères spécifiques, la variabilité de celle-ci soutient la théorie qu’il n’y a qu’une seule espèce Homo avec une variabilité inter et intra-populations." Lordkipanidze propose que ces individus représentent une espèce apparue peu après la transition entre l'Homo habilis et l'Homo erectus, il y a environ 1.8 million d'années. Mais il préfère ne pas les assigner à une nouvelle espèce humaine. Sans évoquer une spéciation, il s'agirait plutôt d'une simple adaptation locale et donc d'une sous-espèce de l'Homo erectus. Comme d'autres l'ont fait avec "l'homme de Pékin", compte tenu de la variabilité observée, aujourd'hui la majorité des chercheurs considère que les fossiles humains de Dmanisi appartiennent à la sous-espèce Homo erectus georgicus. L'aventure dans les gènes La découverte de ces ossements en Eurasie remontant à 1.8 million d'années a conduit les chercheurs à revoir leurs connaissances sur la migration des premiers hommes en dehors de l'Afrique, indépendamment de savoir par où, quand et comment ils arrivèrent en Géorgie ou en d'autres lieux du globe.
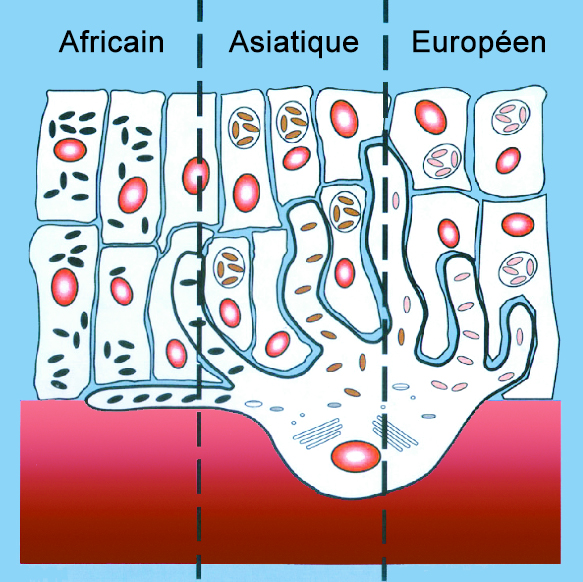
On ignore de quelle région d'Afrique sont originaires les individus de Damnisi ou leurs ancêtres. Quand on examine la carte du monde présentée à gauche, plus de 3500 km séparent le site de Dmanisi du nord de l'Egypte et plus de 5000 km le sépare du Kenya. S'ils ont pu effectuer ce voyage en quelques années en s'accommodant de quelques périodes de repos, on peut aussi imaginer qu'ils rejoignirent la Géorgie par hasard, au terme de plusieurs générations d'une vie de nomadisme à travers les contrées encore vierges de l'Afrique du Nord et du Moyen-Orient où la vie était littéralement florissante. En effet, à cette époque du Calabrien, le climat en Afrique de l'Est et du Nord ainsi qu'au Moyen-Orient était beaucoup plus humide qu'aujourd'hui et la faune tropicale du Villafranchien très riche et diversifiée, alors que les régions tempérées et nordiques subissaient un âge glaciaire de plus. Il y a encore quelques années, les chercheurs pensaient que pour conquérir le monde, nos ancêtres devaient remplir des conditions particulières, hors de portée des premiers humains, notamment de l'Homos erectus. Ils pensaient qu'il fallait avoir un grand cerveau, preuve d'intelligence, être grand pour mieux apprécier les dangers et vaincre les obstacles, être équipé d'armes et d'outils sophistiqués pour survivre aux aléas du voyage et avoir beaucoup d'audace et d'esprit d'aventure pour se lancer le défi de partir à la découverte du monde inconnu situé par delà l'horizon. Encore aujourd'hui, pour certaines personnes, même curieuses de nature, il est très inconfortable et angoissant de partir à l'aventure en dehors des chemins balisés. On peut imaginer tout l'angoisse et l'inquiétude que devaient ressentir des humains primitifs explorant des contrées sauvages, même si la vie nomade avec ses risques inhérents faisait partie de leur quotidien. Ne parlons même pas de la peur et de l'angoisse qu'ils devaient ressentir la nuit, blottis au fond d'une grotte plongée dans l'obscurité totale quand ils entendaient le bruit des bêtes sauvages rôder autour d'eux. Or les squelettes de Dmanisi prouvent que les hommes ont quitté l'Afrique plus tôt qu'on le pensait, sous une forme encore primitive et sans disposer nécessairement d'outils sophistiqués. Emportant avec eux le strict nécessaire, leur volonté et leur confiance dans leur savoir-faire ont convaincu cette petite tribu que cette odyssée était à leur portée. Tout le symbole de l'ambition de l'humanité. Comme nous l'avons déjà évoqué, cette idée de partir en expédition vers des terres inconnues ne fut pas étrangère à l'Homo erectus dont on retrouve les traces bien loin en dehors de l'Afrique. Cette idée de toujours vouloir partir pour découvrir d'autres terres et d'autres peuples accompagna les hommes durant plusieurs millions d'années, jusqu'à ce qu'ils décident de se sédentariser. Mais notre histoire nous démontre que cette curiosité et ce besoin de connaissance ou simplement d'évasion est pour ainsi dire empreint dans nos gènes. Variabilité de la pigmentation de la peau : au moins 900000 ans Alors que certaines personnes se sont tournées vers la science pour soutenir l'origine biologique du concept de race et que certains mouvements d'extrême droite (néonazis, suprémacistes, eugénistes, etc.) l'instrumentalisent pour nous faire croire à la suprématie de la race blanche et son intelligence supérieure, depuis quelques décennies la recherche a démontré que la génétique n'avait pas grand chose à voir dans la pigmentation de la peau et a démontré qu'en ce qui concerne l'être humain, l'idée qu'il existerait différentes races est une ineptie.
On sait depuis longtemps que chez l'homme les gènes de la pigmentation sont distribués en différents endroits des chromosomes et qu'il n'y a pas de gène de la pigmentation sur le chromosome sexuel X. Il s'agit des gènes SLC24A5, SLC45A2 (ou MATP), OCA2, DCT, TYRP1, LYST, MITF, MFSD12, ADAM17 et ADAMTS20 qui interviennent dans la mélanogénèse ainsi que le gène blond KTLG. Il faut également citer MCR1 principalement présent chez les Caucasiens (cf. Les Mélanésiens blonds). Certains gènes comme SLC24A5 présentent une allèle mutante (Phe374Leu) qui est pratiquement toujours présente dans les populations européennes et quasiment absente ailleurs et notamment en Afrique centrale comme l'ont montré les études sur la pigmentation de G.Barsh (2003) et Esteban Parra (2007). Ces mêmes études montrent que certains parmi ces gènes sont uniques à une population tandis que d'autres sont distribués à travers plusieurs ou même l'ensemble des populations. Ainsi certains gènes de la pigmentation sont uniquement présents chez les Européens (SLC45A2 ou MATP), chez les Eurasiens (ADAM17, ADAMTS20) ou chez les Africains (MFSD12), d'autres uniquement chez les Européens et les peuples du Moyen-Orient (SLC24A5), chez les Européens et les Eurasiens (OCA2, KTLG), chez les Européens et les Africains (TYRP1), chez les Eurasiens et les Africains (DCT, LYST) ou encore chez les Européens, les Eurasiens et les Africains (MITF). La généticienne Sarah Tishkoff de l'Université de Pennsylvanie et son équipe ont publié en 2017 dans la revue "Science" les résultats d'une étude à grande échelle démontrant que les peaux claire et sombre ont coexisté pendant des centaines de milliers d'années. L'équipe composée de 47 chercheurs multidisciplinaires spécialisés dans les sciences de la vie (génétique, biologie, médecine, épidémiologie, hématologie, oncologie, physiologie, etc) mais également en paléontologie, mathématiques et statistique a mesuré la pigmentation cutanée de plus de 2000 personnes génétiquement et ethniquement diverses originaires de Tanzanie, d'Éthiopie et du Botswana. Les chercheurs ont analysé le génome de 1570 d'entre elles et identifié huit sites sur les chromosomes associés à la pigmentation de la peau dont le gène SLC24A5 et sa variante précitée.
Une hypothèse à laquelle les chercheurs ont longtemps souscrite concernant l'évolution de la couleur de la peau était que l'Homo sapiens qui vivait originellement en Afrique présentait une peau sombre, fortement pigmentée de mélanine pour se protéger du rayonnement ultraviolet intense du Soleil. Les humains ayant ensuite quitté l'Afrique, on pensait que les mutations conduisirent à une peau plus claire capable de réguler la production de vitamine D. Pour rappel, chez l'homme 80% de la vitamine D est synthétisée à partir d'un dérivé du cholestérol appelé cholécalciférol qui synthétise la vitamine D3 sous l’action des rayonnements UV-B du Soleil (d'où son surnom de "vitamine soleil"). Elle intervient dans l’absorption du calcium et du phosphore par les intestins et sa réabsorption résiduelle par les reins. Elle est donc essentielle à la croissance et la robustesse du squelette. Mais la nouvelle étude montre que l'évolution de la couleur de la peau est beaucoup plus complexe. Chacun des sites de l'ADN impliqué dans la pigmentation présente des variantes associées à la fois à la peau claire et la peau sombre. Sept variantes sont associées à une peau claire sont apparues il y a plus de 270000 ans et quatre autres variantes remontent à plus de 900000 ans. Sachant que l'Homo sapiens n'a pas évolué jusqu'il y a 200000 à 300000 ans, cette découverte suggère que les gènes responsables des peaux claires étaient présents dans le matériel génétique de nos ancêtres préhumains des centaines de milliers d'années avant que le premier humain (Homo) ne marche sur la Terre. Notons que les populations africaines à peau noire présentent un très faible polymorphisme du gène MC1R - l'un de ceux qui contrôlent la couleur des cheveux et la pigmentation - contrairement à ce qu'on trouve chez beaucoup d'autres gènes. Cela signifie que les allèles variants susceptibles d'entraîner une nette diminution de la pigmentation n'ont pas réussi à s'implanter dans ces populations, suggérant qu'ils furent éliminés par la sélection naturelle (probablement du fait qu'il s'agit d'un gène récessif - l'enfant l'hérite des deux parents - et qu'il fut affecté par le stress engendré par les UV solaires sur la peau). En parallèle, nous verrons que certains Homo sapiens européens comme l'homme de Cheddar découvert dans le Somerset en Angleterre ont conservé une peau sombre jusqu'il y a 10000 ans mais avaient déjà probablement les yeux clairs. Les études suggèrent que les gènes de la pigmentation sont plus mobiles qu'on le pensait. Trois des gènes associés à la peau plus foncée ont probablement évolué à partir de gènes des peaux plus claires, ce qui signifie que les personnes ayant une peau sombre (par exemple les bergers du Sahara) ont développé cette pigmentation à une époque récente. Si beaucoup de chercheurs pensaient que seule la peau claire évoluait, selon Tishkoff "la peau foncée continue aussi d'évoluer". Ces résultats ajoutent une composante inattendue à l'histoire récente de la couleur de la peau. Ainsi, les personnes à la peau sombre du sud de l'Inde, de l'Australie et de la Nouvelle-Guinée n'ont pas subi une évolution pigmentaire de façon indépendante, simplement parce que l'évolution le favorisa. En réalité, ils ont hérité des variantes génétiques sombres ancestrales que l'équipe de Tishkoff découvrit chez les Africains. Concernant le gène SLC24A5, l'étude montre également que sa variante associée à la peau claire partagée entre les Européens et les peuples du Moyen-Orient s'est déployée au cours des derniers milliers d'années, retournant même en Afrique pendant les vagues de migration du Moyen-Orient.
Quant au concept de race suranné depuis longtemps, Tishkoff confirme à son tour qu'il s'agit d'une construction arbitraire et sociétale qui n'a rien à voir avec la génétique. Il y a également une variation continue de la couleur de la peau sombre. Selon Tishkoff, classer les humains selon leur couleur de peau est une terrible erreur : "l'étude discréditait vraiment l'idée d'une construction biologique de la race : il n'y a pas de frontières distinctes parmi les groupes compatibles avec les marqueurs biologiques." En résumé, les gènes de la peau claire existent depuis très longtemps, bien avant l'apparition des premiers homininés. "Si vous deviez raser un chimpanzé, vous verriez que sa peau est claire, légèrement pigmentée" expliqua Tishkoff dans un communiqué de presse. "Donc, il est probable que lorsque nous avons perdu nos poils, nous étions censés être dans la clairière d'une savane ouverte. Les mutations influençant à la fois la peau claire et la peau foncée ont continué à évoluer chez les humains, même au cours des derniers millénaires", conclut-elle. Prochain chapitre
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||





















{kind=link}