|
|
|
La perte de gènes ancestraux comme moteur de l'évolution
Les conséquences d'une perte adaptative de gènes La perte de la fonction de certains gènes peut-elle ou non favoriser l'adaptation des espèces à de nouvelles conditions, voire déclencher une macroévolution ? C'est à cette question qu'ont tenté de répondre le biologiste évolutionniste Matthias Huelsmann de l'Institut Max Planck de Biologie Cellulaire Moléculaire et Génétique (MPI-CGB) et son équipe dans une étude publiée dans la revue "Science" en 2019. On sait que les mutations génétiques constituent le principal moteur de l'évolution. Elles se produisent dans des gènes, créant des variants génétiques qui permettent aux espèces de s'adapter aux changements environnementaux. Ces gènes mutants sont appelés des gènes orthologues quand ils sont les descendants d'une séquence unique. Les duplications de gènes sont une autre manière qu'ont trouvé les espèces pour s'adapter aux changements environnementaux. Les gènes issus d'une duplication sont appelés des gènes paralogues. Ils sont soustraits à la pression sélective exercée sur le gène ancestral. Tout récemment encore, on croyait que l'évolution s'était réalisée essentiellement grâce à l'acquisition de nouveaux gènes orthologues et paralogues, notamment par croisements. Mais dans une étude publiée en 2016 dans la "National Review of Genetics", R.Albalat et C.Cañestro ont montré que la perte de gènes ancestraux constitue aussi un important moteur de l'évolution. Matthias Huelsmann et son équipe ont entrepris de déterminer si la perte de fonction de gènes avait joué un rôle dans la macroévolution des cétacés (baleines et dauphins). Les cétacés et les hippopotames modernes ont divergé et évolué à partir d'un ancêtre commun terrestre qui vivait durant l'Eocène il y a environ 50 millions d'années. Alors que les hippopotames se sont adaptés à une vie semi-aquatique, les cétacés ont acquis durant une phase de transition la plupart des traits anatomiques et physiologiques qui leur ont permis de s'adapter à une vie exclusivement aquatique : corps fuselé, peau épaisse et bien isolée, système respiratoire performant, grande capacité de stockage de l'oxygène, perte des membres postérieurs, etc. Ces adaptations ont certainement nécessité l'acquisition de nouveaux gènes mais la perte de gènes a-t-elle aussi été nécessaire ? Pour le savoir, les chercheurs ont examiné les séquences génomiques de 62 espèces de mammifères : 55 mammifères terrestres, 3 cétacés odontocètes (cétacés à dents : dauphin, orque, cachalot), 1 cétacé mysticète (rorqual), 2 pinnipèdes (phoque, morse) et 1 lamantin. Ils les ont comparées au génome de référence humain GRCh38 en se concentrant sur 19769 gènes humains. Il s'agissait de détecter parmi ces milliers de gènes ceux qui auraient été perdus chez les cétacés ancestraux, plus particulièrement au cours de la transition ayant conduit les ancêtres des cétacés modernes vers la vie totalement aquatique. Mais comment détecter la perte de certains gènes ou plutôt la fonction des gènes concernés ? Les chercheurs devaient identifier parmi ces 19769 gènes ceux portant une ou des mutations susceptibles d'entraîner la perte de la fonction de la protéine correspondante. Ainsi, la présence d'un triplet TAG dans un exon sur la chaîne sens d'ADN (ou ADN de codage), celle qui est utilisée pour la comparaison, entraîne l'apparition d'un codon anti-sens (ou brin de matrice) UAG à l'intérieur de l'ARN messager (ARNm). Sa présence interrompt prématurément la traduction, ce qui a pour résultat de produit une protéine incomplète et donc inactive. La détection d'une insertion ou d'une délétion (la perte d'un fragment d'ADN) d'un ou plusieurs nucléotides (mais pas un multiple des 3 lettres choisies parmi A, U, G et C) dans l'ADN sens (mutations frameshifts) change le cadre de lecture de l'ARNm. Tous les codons en aval de la mutation sur ce dernier sont modifiés, ce qui entraîne l'incorporation d'autant de mauvais acides aminés dans la protéine correspondante et inactive cette dernière. La délétion d'un ou plusieurs exons dans l'ADN entraîne également la perte de plusieurs acides aminés et la production d'une protéine non fonctionnelle. Une mutation dans l'ADN au niveau d'une séquence d'excision-épissage de l'ARN prémessager perturbe la maturation de ce dernier et produit une séquence anormale d'exons sur l'ARNm et une protéine inactive. Les auteurs ont identifié 236 gènes humains dont les orthologues chez les cétacés ont subi une mutation. Permi ceux-ci, 3 gènes ont été observés chez les 55 mammifères terrestres étudiés, ce qui montrent que ces gènes sont impliqués dans l'adaptation à la vie aquatique. 110 gènes portent des mutations inactivatrices qu'on trouve à la fois chez les odontocètes et les mysticètes ce qui suggère qu'elles sont apparues avant la divergence de ces deux clades de cétacés (voir cladogramme ci-dessous). Finalement, en comparant les 110 gènes humains de référence aux séquences génomiques des hippopotames, les chercheurs ont montré que 85 gènes n'ont pas varié chez les hippopotamidés. Ces gènes sont donc ceux qui ont été inactivés soit durant la transition il y a 50 millions d'années de la vie terrestre à la vie aquatique chez les ancêtres des cétacés soit après la divergence des hippopotamidés des cétacés mais avant celle des deux clades de cétacés odontocètes et mysticètes.
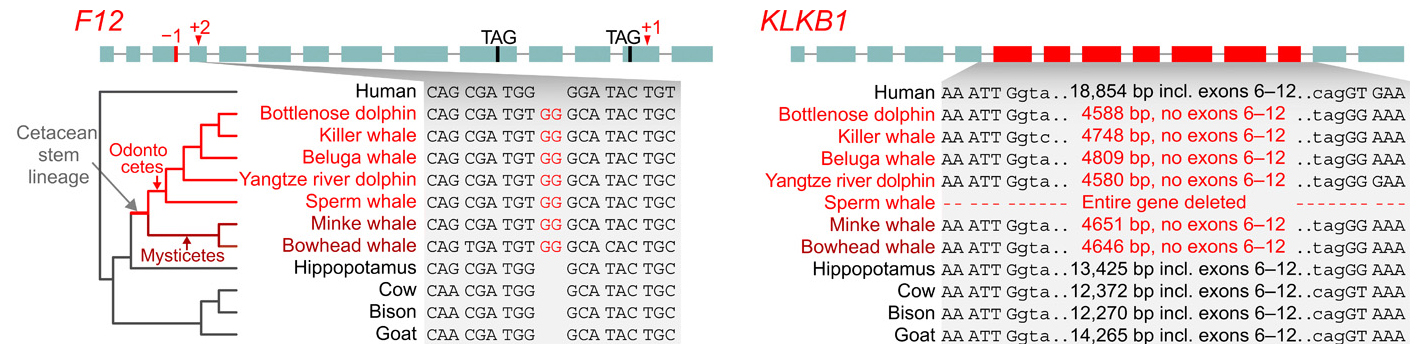
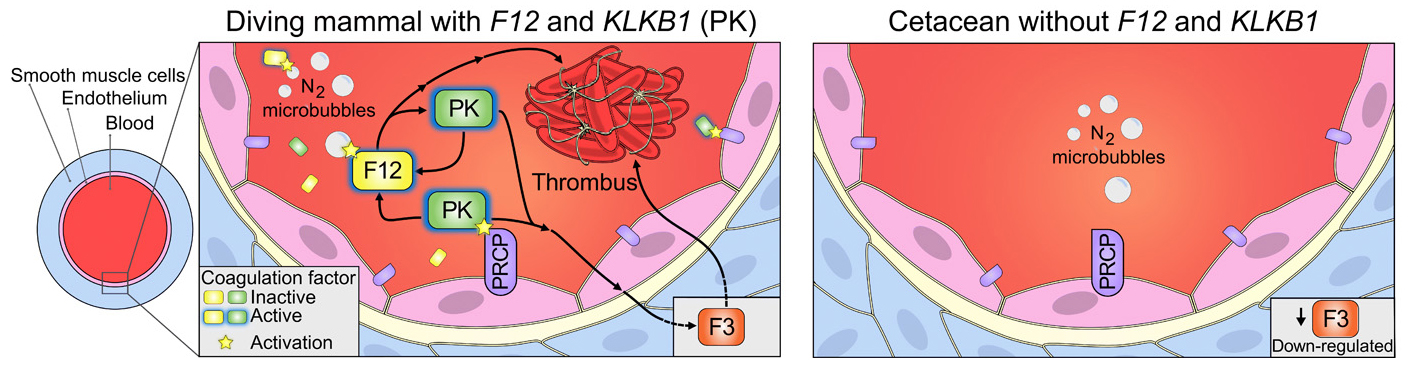
Selon les auteurs, la perte de gènes au cours de l'évolution ne résulte pas nécessairement d'un relâchement de la sélection naturelle sur une fonction devenue inutile mais elle peut constituer un véritable mécanisme d'adaptation. Mais comment ces pertes de fonctions génétiques chez les cétacés ont-elles pu favoriser leur adaptation ? La réponse à cette question exige de connaître la fonction normale de ces gènes. En observant le rôle des gènes orthologues chez l'être humain et les effets de leur inactivation provoquée chez la souris, il est possible de savoir si leur perte chez les ancêtres des cétacés il y a 50 millions d'années a constitué ou non un avantage sélectif dans la conquête du milieu aquatique. Pour cela les chercheurs ont étudié trois des 85 gènes identifiés : F12, KLKB1 et POLM (ADN polymérase mu). Selon les chercheurs, les facteurs de coagulation XII (F12) et KLKB1 (kallikréine B1) ont été perdus dans la lignée des cétacés, ce qui correspond aux constatations des études antérieures. Les cladogrammes illustrent les exons codants superposés aux mutations inactivantes des gènes communs aux odontocètes et aux mysticètes. Ces mutations sont donc probablement survenues avant la divergeance de ces lignées. Les cladogramme présentés ci-dessus illustrent l'évolution de ces deux gènes : F12 et KLKB1. Les exons (segments d'un précuseur ARN) sont représentés par les boîtes vertes. La transcription par l'ARN polymérisase II se réalise de gauche à droite. Les chiffres en rouge (-1, +2 et +1) au-dessus de l'ADN de F12 indiquent la position des mutations dites "frameshifts". Les boîtes en rouge dans l'ADN de KLKB1 représentent la délétion des exons 6 à 12 de ce gène. Les triplets TAG correspondent aux mutations anti-sens qui provoquent l'arrêt de la traduction. On constate que F12 a accumulé plusieurs mutations inactivatrices distinctes. La mutation +2 (équivalente à une mutation -1) dans l'exon 4 de F12 correspond à l'insertion de deux nucléotides G (plus précisément deux dGMP) dans la séquence de l'exon 4 (en gris) de tous les cétacés étudiés. Cette mutation est absente chez les mammifères terrestres. À elle seule, elle a inactivé la protéine en changeant le cadre de lecture de l'ARNm et la séquence de tous les acides aminés situés en aval de son site (non illustré). Le cladogramme présenté à droite montre que la délétion des exons 6 à 12 codant pour KLKB1 (en rouge) s'est produite dans la lignée des cétacés (les bases introniques adjacentes aux exons 5 et 13 sont en minuscules). Dans le cladogramme de gauche, F12 code pour un zymogène (le précurseur protéique d'une enzyme) qui s'active automatiquement au contact de diverses surfaces, notamment des microbulles d'azote qui se forment lors de plongées en apnée. Dans le cas de KLKB1, la délétion des 7 exons entraîna l'inactivation de la protéine en éliminant de nombreux acides aminés. Elle s'est aussi produite spécifiquement chez les cétacés. Chez l'être humain, les gènes F12 et KLKB1 codent chacun pour un facteur inactif de coagulation, plus précisément un précurseur de protéase (enzyme qui clivent des protéines). Au simple contact dans le sang avec une surface biologique, le précurseur produit par F12 change de conformation et devient une protéase active (voir schéma ci-dessous). Celle-ci peut alors cliver le précurseur de KLKB1 et le convertir en une autre protéase active, la kallikrein (désignée pour simplifier par PK). PK agit sur une protéine cible spécifique et entraîne indirectement la production du facteur de coagulation F3. Ce dernier active les plaquettes sanguines et provoque leur agglutination et la formation d'un caillot de sang. Le produit inactif de KLKB1 peut aussi être activé par une autre enzyme, la protéase PRCP, liée celle-là à l'endothélium des vaisseaux sanguins. Lors de leurs fréquentes plongées en apnée, les cétacés subissent un ralentissement cardiaque et une vasoconstriction de leurs vaisseaux, ce qui réduit la circulation sanguine périphérique. Il y a aussi formation de microtubules dans le sang. Sans l'inactivation de F12, la protéase inactive produite par ce gène aurait de fortes chances de s'activer par contact avec les microtubules du plasma et de provoquer des thromboses. De même sans l'inactivation du gène KLKB1, l'activation de son produit en une protéase PK serait favorisée par la constriction des vaisseaux qui rapprocherait ce produit de PRCP, ce qui augmenterait le risques de thromboses. L'inactivation des gènes F12 et KLKB1 chez les cétacés semble donc le résultat d'une adaptation bénéfique. Notons que les gènes F12 et KLKB1 ont déjà été inactivés chez la souris par transgenèse in vivo. On a constaté qu'une telle invalidation génique évite l'apparition de thromboses provoquées mais n'empêche pas la guérison des blessures.
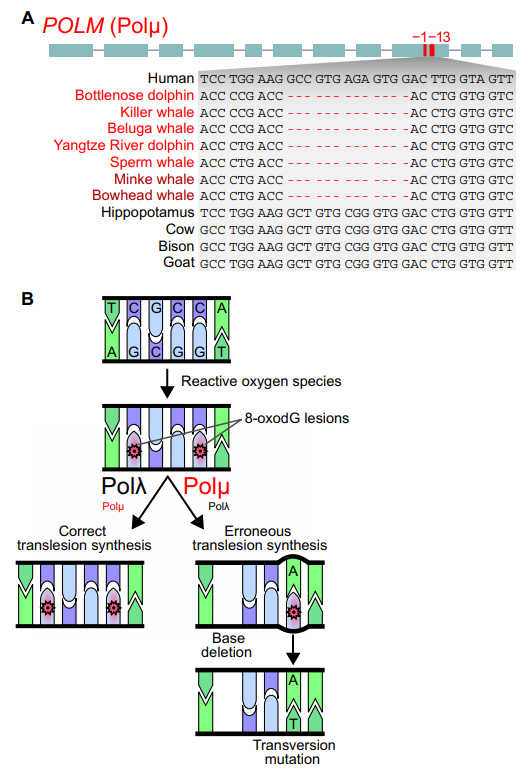
Autre exemple avec le gène POLM. Son inactivation chez les cétacés semble aussi avoir contribué à leur adaptation et leur macroévolution. Diverses mutations observées chez tous les cétacés étudiés ont été décelées dans ce gène, dont notamment la délétion de 13 nucléotides dans l'exon 9. Les ancêtres des cétacés ont dû contourner le problème de la formation d'espèces réactives à l'oxygène (ROS) dans le sang lorsque la diminution de l'afflux sanguin (l'ischémie) dû à la vasoconstriction des vaisseaux périphériques est suivie d'un rétablissement de la circulation sanguine. Ces ROS créent des lésions dans l'ADN en modifiant des bases, comme l'oxydation de la guanine (en 8-oxodG). Les ROS peuvent égalemen briser les doubles brins d'ADN. La réparation de ces cassures est principalement assurée par la protéine Ku70/80 qui intervient dans un mécanisme de jonction d'extrémités non homologues (ou NHEK, Non-Homologous End-Joining). Au cours de cette réparation, deux autres enzymes de réparation de l'ADN, Polm lambda et Polm mu, interviennent pour combler les régions simple brin restantes. Polm mu est plus efficace que Polm lambda pour agir sur divers types de cassures mais son activité polymérasique est sujette à des erreurs. Elle provoque des délétions de bases et peut incorporer le mauvais nucléoside devant l'ADN modèle, ce qui engendre des mutations. L'inactivation du gène POLM chez les cétacés semble donc présenté l'avantage de réduire les conséquences néfastes sur l'ADN du stress oxydatif associé aux plongées en apnée en réservant à Polm lambda la tâche d'assurer une réparation plus fidèle des cassures dans l'ADN. La fonction de plusieurs autres gènes humains orthologues d'autres gènes des cétacés (parmi les 85 gènes inactivés) montre aussi que la perte de ces gènes n'est pas la conséquence d'un relâchement de la sélection naturelle mais au contraire le résultat d'une adaptation.
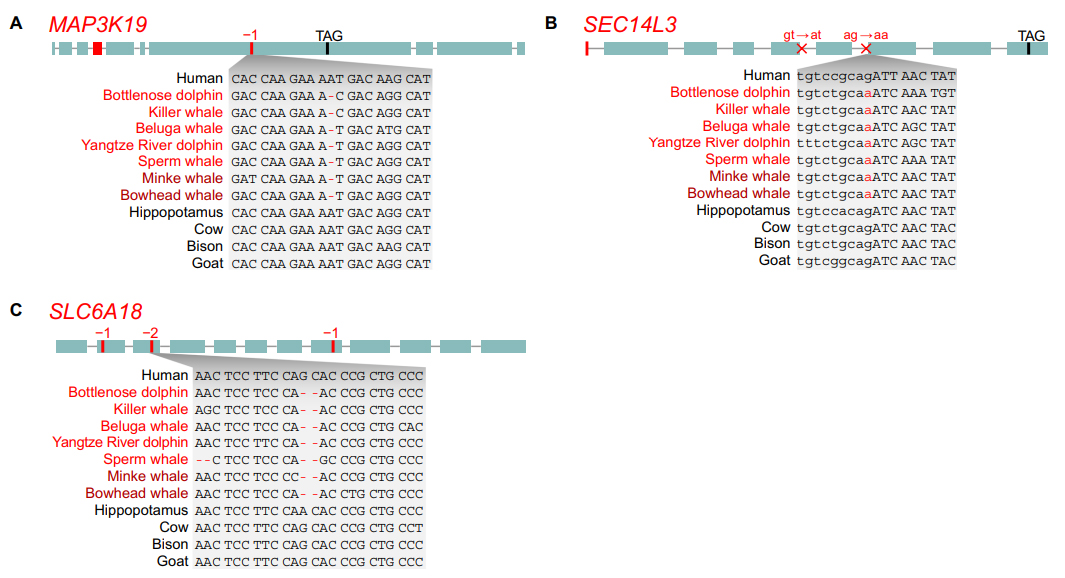
En guise de conclusion La transition de la terre à l'eau chez les baleines et les dauphins (cétacés) s'est accompagnée d'adaptations génétiques et physiologiques remarquables. Les chercheurs ont trouvé 85 pertes de gènes. Certains d'entre eux étaient probablement bénéfiques pour les cétacés, par exemple en réduisant le risque de formation de caillots de sang pendant la plongée (F12 et KLKB1), réparant les dommages de l'ADN erroné (POLM) et une inflammation des poumons induite par le stress oxydatif (MAP3K19). Des pertes génétiques supplémentaires peuvent refléter d'autres adaptations liées à la plongée, telles qu'une vasoconstriction améliorée au cours de la plongée (SLC6A18) et la modification de la composition tensioactive pulmonaire (SEC14L3), tandis que la perte de SLC4A9 est liée à un besoin réduit de salive. Enfin, la perte de la synthèse de la mélatonine et des gènes du récepteur (AANAT, ASMT et MTNR1A/B) peut avoir été une condition préalable à l'adoption du sommeil unihémisphérique. Ces résultats suggèrent que certains gènes perdus chez les cétacés ancestraux ont probablement été impliqués dans l'adaptation à un environnement entièrement aquatique. |
||||||||||||||||||||||||