|
|
|
Les techniques de recherches paléontologiques
Datation des fossiles, filiation génétique et analyses paléontologiques (I) Dans son éternelle quête de connaissance, l'homme a toujours voulu comprendre quel est sa place dans l'Univers, sa structure, son histoire et son évolution ainsi que celles des organismes qu'il cotoie ou découvre en creusant le sol. Cette passion donna naissance à la paléontologie fondée en 1822 par le zoologiste français Henri Ducrotay de Blainville et qui s'est répandue à travers le monde grâce au géologue britannique Charles Lyell, ami proche de Charles Darwin. Son sujet étant tellement vaste et complexe qu'elle englobe de nombreuses disciplines et branches des sciences du vivant aux géosciences, réservant une place de choix à l'étude des ancêtres de l'homme et leur évolution, la paléoanthropologie ainsi qu'aux paléoenvironnements dans lesquels ils vécurent. Grâce à des études multidisciplinaires (ou interdisciplinaires), dans le meilleur des cas la paléontologie permet d'avoir une très bonne estimation de l'âge des fossiles et de la biocénose, de l'aspect de l'écosystème, des conditions climatiques et de l'impact de l'homme sur l'environnement. En effet, au terme de plusieurs années voire dizaines d'années de recherches collectives et de tentatives d'interprétations, si tout se passe bien les scientifiques ont accumulé tellement de connaissances et de résultats dans tous les domaines qu'ils peuvent retracer l'histoire du site et de ses occupants à l'époque où ils étaient actifs y compris, pour les périodes préhistoriques, sur le plan socio-économique. Ainsi, parfois c'est un pan de la vie de toute une tribu ou de la population d'un village qui revit grâce au travail méticuleux des chercheurs, des modélisations et des simulations informatiques. Voyons plus en détails les méthodes de travail de ces chercheurs. La recherche en paléontologie comprend cinq grandes étapes : la découverte des fossiles, l'extraction, la datation, l'identification et l'interprétation. Tous les fossiles n'affleurent pas à la surface du sable comme au Tchad ou sur certains versants des gorges de l'Olduvai. Si la découverte paraît fortuite aux yeux du grand public, pour les paléontologues ou les archéologues pour citer deux professions, elle résulte surtout des résultats des études réalisées en amont et d'un calcul de probabilité; on ne fouille pas au hasard mais dans des sites préalablement sondés susceptibles de contenir des échantillons ou proches de gisements existants. Bien que les travaux d'excavations soient généralement assez rapides (quelques mois parfois étalés sur plusieurs saisons de fouilles), l'extraction complète des vestiges peut parfois durer plusieurs années quand le gisement est étendu ou lorsque le fossile est littéralement soudé dans une gange minéralisée aussi dure que du béton qu'il faut retirer très méticuleusement millimètre par millimètre. En attendant, les chercheurs peuvent déjà travailler sur des moulages et des copies conformes ou à partir de modèles numérisés. Concernant l'identification du spécimen, dans certains cas elle est tout simplement impossible à réaliser du fait que le spécimen est réduit à quelques fragments et l'espèce unique en son genre. C'est à peine si on peut déterminer s'il s'agit par exemple d'une espèce Australopithèque ou Homo. Parfois, il faut patienter des années jusqu'à ce que par chance d'autres fossiles de la même espèce, plus complets ou mieux conservés permettent de rapprocher l'holotype avec les nouveaux échantillons. C'est ainsi qu'il s'est écoulé plus de 40 ans avant que certains ossements d'Homo habilis et d'Homo rudolfensis soient correctement assignés à leur propiétaire respectif. Parmi toutes ces activités, la datation des échantillons est encore le processus le plus rapide. Il repose sur plusieurs techniques, souvent complémentaires selon la qualité, la nature ou la taille de l'échantillon, parmi lesquelles : - la radioactivité du radiocarbone ou carbone-14 ou d'autres éléments - la structure temporelle des populations (TPS) - l'horloge moléculaire de l'ADN mitochondrial - la thermoluminescence - la résonance de spin électronique (ESR) - la luminescence optiquement simulée (OSL) - le paléomagnétisme - la dendrochronologie - la racémisation des acides aminés - la typologie - la biochronologie. En général, l'interprétation des données ne se limite pas aux études purement paléontologiques. Les chercheurs s'appuient beaucoup sur l'étude des paléo-écosystèmes (biotope et biocénose ancestraux) et du paléoclimat qui font elles-mêmes généralement appel aux géosciences et aux sciences du vivant. Parmi ces sciences, disciplines ou branches qui viennent en support de la paléontologie, citons (la liste n'est pas exhaustive) la paléobiologie, la paléoclimatologie, la protistologie, la physiologie, la biochronologie, la phylogénie, la paléogénétique, l'archéologie, la géoarchéologie, l'archéobiologie, la paléozoologie, la paléobotanique, la géologie, la pédologie, la micropaléontologie, la micromorphologie, l'hydrogéologie, l'océanographie, la dendrochronologie, la sclérochronologie, la glaciologie, la minéralogie. Nous définirons chacune d'entre elles page suivante. Pour les périodes allant de la Préhistoire à l'époque contemporaine, les chercheurs peuvent également affiner leurs résultats sur les plans morphologiques, socio-économiques, politiques et migratoires notamment en recoupant ou complétant ces données avec les résultats obtenus par les recherches archéologiques, anthropologiques, génétiques, linguistiques et le travail des historiens. Voyons à présent les différentes techniques de datation des fossiles et des artefacts avant de décrire les différentes sciences et disciplines précitées. Techniques de datation des fossiles et des artefacts Radioactivité du carbone-14 La méthode de datation la plus connue est la datation au carbone-14 qui est utilisée dans de nombreuses disciplines des sciences de la Terre et du vivant. C'est une méthode de datation absolue qui permet de fixer l'artefact chronologiquement dans le temps.
La méthode est basée sur la désintégration d'un nucléide radioactif. Le noyau père instable du 14C a une demi-vie ou période de 5730 ±40 ans. Autrement dit, au bout d'environ 5730 ans, 50% des isotopes de 14C seront transformés en 12C stable. 5730 ans plus tard, 50% des isotopes restants de 14C seront transformés en 12C et ainsi de suite. Il arrive donc un jour où l'objet ne contient pratiquement plus de 14C mais uniquement du carbone 12C et 13C. La mesure de la teneur en radiocarbone n'est possible que sur des matières d'origine organique. La raison est liée au fait qu'un organisme vivant assimile le carbone sans distinction isotopique; la proportion de 14C par rapport au carbone total (12C + 13C + 14C) restant identique à celle existant dans l'atmosphère à l'époque où vécut l'organisme. Dès l'instant où l'organisme meurt, la proportion de radiocarbone qu'il contient commence à décroître selon la loi exponentielle de désintégration (ou de décroissance) du 14C. Ainsi, à partir d'un échantillon de matière organique on peut mesurer le rapport 14C/Ctotal et en déduire son âge. Les mesures du rapport isotopique sont réalisées par spectrométrie de masse (et plus rarement par thermoluminescence ou par luminescence stipulée optiquement ou par infrarouge) qui permet de dater de très petits échantillons (< 1 mg) en moins d'une heure. Bien qu'utilisant un accélérateur de particules, les installations les plus petites occupent à peine 2 mètres cubes, rendant cette technologie accessible à de petits laboratoires d'analyses. De façon générale, l'âge Δt de l'échantillon obéit à la relation suivante : Δt = ln (a+1) / λ avec, Δt = t - to, où t est l'instant de la mesure et to l'instant de la mort de l'organisme ln, le logarithme népérien a, le coefficient directeur de la droite de régression soit Co/Ctotal, où Co est la quantité de carbone à l'instant to et Ctotal = 12C + 13C + 14C λ = 1.210x10-4 an-1 est la constante de décroissance du 14C Par exemple, pour a = 50.6, Δt = 32591 années. Lorsque la mesure est effectuée, l'âge est exprimé en années "before present" (BP) ou âge conventionnel. Ce nombre absolu doit ensuite être corrigé pour tenir compte de la présence récente de carbone fossile et de 14C dans l'atmosphère (cf. U.Lyon) et par rapport à une date de référence pour finalement aboutir à l'âge "calibré" réel, celui que tous les chercheurs utilisent dans leurs publications. Etant donné que la période du radiocarbone est d'environ 5730 ans, son utilisation devient difficile au-delà de 40 à 50000 ans. Pour les datations plus anciennes, il faut donc utiliser d'autres éléments instables dont la période de désintégration est connue comme par exemple le couple 234U-230Th dont la période du 230Th est de 65200 ans. On peut ainsi dater des échantillons de carbonates (la calcite supportant les peintures rupestres, les os, les coquillages, les foraminifères, les coraux, les stalagmites et stalactites, etc) remontant jusqu'à 500000 ans. Pour les artefacts très anciens, généralement on utilise des nucléides radiogènes dont la période est de plusieurs millions ou milliards d'années comme le couple 40K-40Ar (qui exploite la méthode Ar-Ar) dont la période du 40K est de 1.25 milliard d'années ou des radioculéides plus lourds comme le couple 87Rb-87Sr dont la période du 87Rb est 48.8 milliards d'années. Comme la plupart des méthodes de datation, toutes ces techniques sont complexes et demandent un traitement de l'échantillon et une analyse en laboratoire. On reviendra sur l'utilisation des systèmes isotopiques à propos de la datation des météorites. A
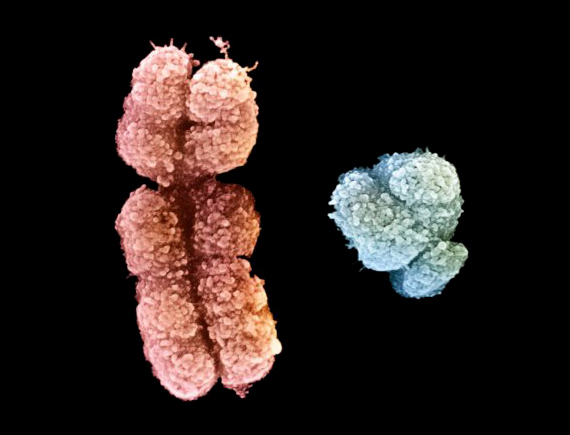
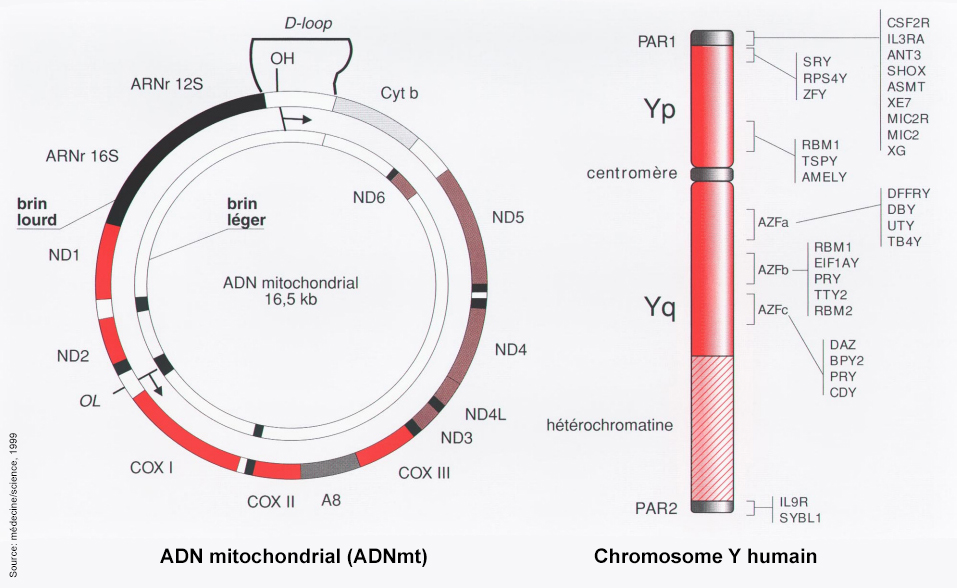
voir : Le temps et les roches La spectrométrie de masse - Le spectromètre de masse par accélérateur La TPS (Time Population Structure) La technique dite TPS (Time Population Structure) ou Structure Temporelle des Populations est une nouvelle méthode de datation absolue des squelettes proposée en 2018 par le chercheur postdoctorant Umberto Esposito du laboratoire du Dr Eran Elhaik de l'Université de Sheffield. La TPS utilise le taux de mutations génomique pour déterminer la date d'un échantillon biologique. Selon l'auteur, la TPS peut calculer les mélanges d'ADN provenant de périodes différentes et estimer l'âge définitif. TPS a déjà montré que ses résultats sont très similaires à ceux obtenus avec la datation traditionnelle au radiocarbone. La différence moyenne calculée sur des échantillons remontant à 45 000 ans entre les prédictions calculées avec TPS et celles datées au radiocarbone était de 800 ans. L'intérêt de la TPS est d'ajouter une méthode à la panoplie existante car elle s'avère utile lorsque par exemple la quantité de radiocarbone est insuffisante pour dater l'échantillon. La technique peut également être utilisée pour caractériser la généalogie des populations ou pour déterminer l'homogénéité des populations dans le cadre d'une étude épidémiologique. Une autre technique très utilisée pour dater les échantillons biologiques repose sur la biologie moléculaire, ce sont les marqueurs génétiques : l'ADN mitochondrial (ADNmt), le chromosome Y, l'ARN ribosomique (ARNr 16S ou 18S), le cytochrome C ou encore l'hémoglobine. Les mitochondries sont des organites autonomes qui possèdent plusieurs copies de l'ADNmt. Chez l'homme, le nombre de mitochondries présentes dans chaque cellule est très variable, allant de 75 mitochondries dans le spermatozoïde à 100000 mitochondries dans l’ovocyte. Ce qui rend ces marqueurs génétiques uniparentaux intéressants tient au fait que les séquences d'ADN portées par l'ADNmt et la région spécifique du chromosome Y ne recombinent pas lors de la méiose et sont donc transmis sans modifications à la génération suivante. En d'autre terme, l'ADNmt se transmet uniquement de la mère aux enfants par les ovules (transmission matrilinéaire) tandis que le chromosome Y se transmet uniquement de père en fils par les spermatozoïdes (transmission patrilinéaire). Les chercheurs étudient surtout l'ADNmt car les mitochondries du spermatozoïde se dégradent rapidement quand elles pénètrent dans l'ovocyte et sont donc rarement exploitables.
Les séquences de l'ADN mitochondrial subissent environ 10 fois plus de mutations que les gènes de l'ADN nucléaire, ce qui permet d’observer une quantité plus importante de variations polymorphiques chez les individus. Généralement les chercheurs utilisent un taux de mutation de l'ADNmt variant entre 2% et 4% par million d'année. La datation peut donc varier du simple au double. Mais en recoupant le résultat avec la datation au radiocarbone et d'autres techniques et en reportant ces estimations dans un graphique multicritères, les chercheurs peuvent isoler une période avec une précision de quelques milliers d'années pour l'Homo sapiens. Ce taux de mutations de l'ADNmt étant relativement constant au cours de l'évolution des espèces, son utilisation comme "horloge moléculaire" permet de retracer dans le temps et dans l'espace l'évolution d'une espèce du côté maternelle, et en particulier l'origine et la distribution des différentes populations humaines et préhumaines. De la même manière, on peut utiliser cette particularité chez le chromosome Y pour étudier l'origine des espèces y compris les goulots d'étranglement génétique, cette fois-ci du côté des hommes. L'utilisation de cette méthode dépend toutefois de la lignée de l'espèce et de l'époque étudiée et est altérée par des facteurs de biais dont il faut tenir compte au risque d'obtenir des datations erronées. En effet, toutes les "horloges moléculaires" ne tournent pas à la même vitesse, ce qui impose leur calibrage. De plus, le taux de mutation dépend des familles d'organismes étudiés. Les analyses doivent également tenir compte d'effets parasitaires comme par exemple des recombinaisons de l'ADN mitochondrial de l'ovule (ovocyte après maturation) par celui des mitochondries du spermatozoïde et autres substitutions génomiques. Enfin, l'analyse est plus difficile à réaliser sur des Australopithèques âgés de plusieurs millions d'années que chez des Néandertaliens de 50000 ans par exemple du fait que la molécule d'ADNmt finit par se dégrader au bout de 100000 ans dans le meilleur des cas. Comme dans de nombreuses autres disciplines, les experts en paléobiologie moléculaire ne peuvent donc pas simplement interpréter les données brutes pour dater un fossile mais ils doivent également vérifier que les données sont de qualité et fiables, au risque d'aboutir à des conclusions aberrantes, comme c'est déjà arrivé. La thermoluminescence Parmi les nombreuses disciplines de la minéralogie, la thermoluminescence est une méthode absolue qui permet de dater les objets cristallins. La thermoluminescence exploite les rayonnements ionisants présents naturellement dans l'atmosphère, en particulier la radioactivité naturelle. Les radioisotopes s'accumulent dans les minéraux tels que les cristaux contenus dans des matières non biologiques qui ont été chauffées : quartz, feldspath, calcite, fluorite, célestine, cryolite, danburite et sphalérite. Lors de la cuisson d'une poterie par exemple, la structure cristalline libère ses éléments radioactifs et l'horloge de la décroissance est remise à zéro. On peut ensuite calculer de débit total d'irradiation ou plaéodose depuis la remise à zéro en chauffant à nouveau l'échantillon. En comparant ce résultat à la valeur enregistrée sur le site par un dosimètre, on peut calculer la date de cuisson de la poterie et par conséquent dater l'occupation du site. Cette technique permet de dater des artefacts comme de la poterie en terre cuite, de la céramique, des fours artisanaux, des traces d'incendies, des noyaux de bronze, des pierres brûlées, des silex chauffés, des roches volcaniques, des stalagmites, des cratères de météorites, des plages et des dunes de sable notamment. Cette technique permet aux scientifiques de dater des objets et des sites archéologiques remontant à 50000 ans mais on peut encore l'utiliser jusqu'à 800000 ans BP avec une précision de 5 à 20%. Si le site a été fortement exposé à la radioactivité naturelle, la limite est d'environ 500000 ans. Cette technique n'est précise que si l'échantillon n'a pas été porté à haute température et, dans le cas des fours, le résutat ne donne que la date de dernière utilisation. NB. A ne pas confondre avec la fluorescence (cf. la "lumière noire" UV, les bâtons lumineux et la gemmologie). La résonance de spin électronique (ESR) La résonance de spin électronique (ESR) est une méthode de datation absolue similaire à la thermoluminescence décrite ci-dessus, mais en utilisant les propriétés magnétiques d'éléments radiosensibles présents dans les minéraux. Cette méthode nécessite au préalable une dosimétrie de l'artefact pour connaître la dose totale de radiations qu'il a reçu depuis son enfouissement. On utilise surtout l'ESR pour dater l'émail dentaire, les os, les concrétions dans les grottes, les coquillages et les grains de quartz. L'ESR permet de dater des objets remontant à plus d'un million d'années voire même plusieurs millions d'années dans le cas des grains de quartz. La luminescence optiquement simulée (OSL) La datation des matières minérales peut s'effectuer au moyen d'une technique appelée la luminescence optiquement simulée (OSL). Elle permet de déterminer à quelle époque des grains minéraux de quartz présents dans les sédiments et préservés du rayonnement solaire furent exposés pour la dernière fois à la lumière, ce qui permet aux archéologues de savoir combien de temps l'artefact ou les pigments par exemple situés dans la même strate sont restés enterrés.
Le paléomagnétisme Le paléomagnétisme étudie l'orientation magnétique des corps ferromagnétiques (laves, etc) par rapport au pôle Nord magnétique. Cette méthode permet de dater les roches (laves, fossiles, etc) de manière absolue. Au-dessus de la température de Curie, les oxydes de fer telles que la magnétite ne sont pas aimantés et ne sont donc pas sensibles au champ magnétique. Placez une boussole près d'une lave en fusion, même si vous y avez fait fondre un kilo d'aimant, son aiguille pointera toujours vers le Nord magnétique. Ce seuil de température se situe à 585°C pour la magnétite et au-dessus de 900°C pour les basaltes et les gabbros. En dessous de 585°C, la lave se fige et les oxydes de fer qu'elle contient vont adopter une orientation précise par rapport au Nord magnétique, celle qu'ils avaient au moment du refroidissement par rapport au Nord magnétique. C'est ce qu'on appelle le magnétisme originel de la lave. Les déviations du pôle magnétique étant connues des géologues et en particulier des vulcanologues, il suffit ensuite de mesurer l'orientation magnétique de ces laves pour déterminer leur âge au moment de leur refroidissement. Indirectement, à partir de l'orientation magnétique on peut déduire à quel endroit se situait la plaque tectonique sur laquelle se sont déposées ces laves à l'époque donnée. Cette technique permet de remonter jusqu'à plus de 100 millions d'années. Elle permet non seulement de calculer l'âge des fossiles prisonniers des laves et de certaines roches sédimentaires magnétiques mais également d'étudier la tectonique des plaques et la dérive des continents. Ces analyses complètent les études géologiques et biochronologiques parmi d'autres sciences de la terre et du vivant. La dendrochronologie
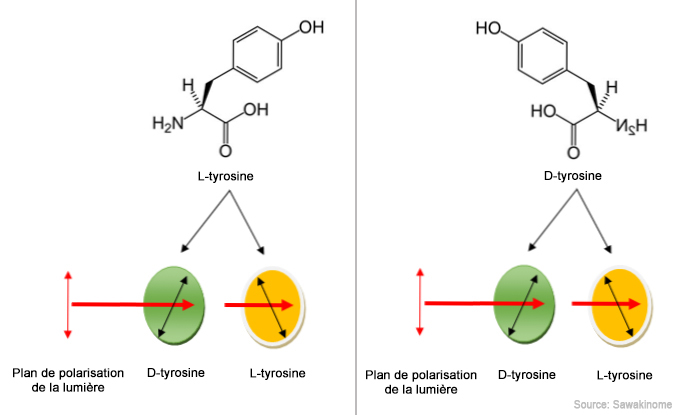
La dendrochronologie est une méthode qui étudie les cernes de croissance des arbres, la cellulose s'imprégnant de l'amplitude des précipitations, de l'activité solaire et des conditions environnementales année après année. Ainsi, certains séquoias de Californie couvrent 8500 ans d'histoire. Les plus grands représentants sur pied mesurent plus de 80 m de hauteur et ont plus de 2000 ans. Cette méthode permet de recueillir des séquences caractéristiques contenant des données scientifiques sur les conditions atmosphériques locales et de dater les arbres. On utilise en général des résineux car ils sont plus sensibles que les feuillus aux variations des conditions de croissance et les feuillus pour étudier les cernes de croissance car leur largeur varie peu. La précision de la datation peut être précise à l'année près. Il s'agit d'une méthode de datation absolue car les séries obtenues peuvent être combinées et mises en correspondance pour obtenir des séquences moyennes qui permettent de dresser une séquence chronologique couvrant des centaines d'années. Cette séquence peut ensuite servir d'étalon de référence. En recoupant les séries de séquences moyennes provenant d'arbres morts à des époques différentes on peut fixer la chronologie dans le temps. On utilise également la dendrochronologie pour calibrer les datations au carbone-14 qui peuvent être imprécises en raison des variations de la concentration en 14C dans l'atmosphère au cours du temps. La racémisation des acides aminés Bien que le titre paraisse pompeux, cette technique fait appel à des concepts relativement simples mais peu connus du grand public qui nécessitent quelques explications. Nous avons expliqué en bioastronomie et en physique des particules que dans la nature beaucoup de phénomènes et de substances privilégient certaines orientations moléculaires et plans de polarisation comme par exemple l'hélice de l'ADN et le dextrose (sucre) qui tournent systématiquement à droite (dextrogyre), jamais dans l'autre sens. C'est la chiralité. En général, si la seconde forme moléculaire ou éniantomère existe, la substance est inactive ou se transforme en poison, bloquant les réactions chimiques de l'organisme qui l'ingère.
La racémisation est la transformation d'une substance optiquement active (polarisée) en un composé dit racémique qui ne dévie pas la lumière polarisée. Cette substance qui présente les deux plans de polarisation et donc les deux éniantomères, gauche et droit, est appelée un stéréoisomère. Il est optiquement inactif, la chiralité laissant place à une racémisation des différentes molécules lévogyres et dextrogyres. Autrement dit, la molécule et son image dans un miroir sont identiques. Les acides aminés sont présents dans toutes les cellules et tous les organismes fondés sur la chimie du carbone. On les retrouve à la fois dans des molécules très simples comme la cytosine (C4H5N3O) constituant l'une des bases de l'ADN et dans les protéines qui sont constituées de chaînes d'acides aminés formant la structure des muscles et des os parmi beaucoup d'autres tissus. Ils sont donc présents en très grande quantité et facile à prélever. Il faut toutefois veiller à ce qu'ils soient bien conservés et purifiés (que les impuretés soient éliminées). La racémisation exploite les propriétés de la chiralité des acides aminés pour dater les organismes fossiles ou gelés dans le permafrost. En effet, pour une raison inconnue mais probablement liée à l'histoire de notre planète, les acides aminés naturels (pas ceux créés en laboratoire) présentent un seul plan de polarisation : ils tournent à gauche (lévogyre). Cependant, après la mort de l'organisme, ce mécanisme de contrôle de la chiralité disparaît et la racémisation reprend ses droits : la proportion des acides aminés lévogyres et dextrogyres s’équilibre. Le processus évolue en fonction de différents facteurs comme la température, l'acidité et des paramètres environnementaux. Concrètement, la vitesse de racémisation varie entre quelques heures dans une eau acide bouillonnante à plusieurs milliards d'années dans une couche sédimentaire sèche et froide. L'étude de la racémisation permet donc de dater les fossiles de manière relative jusqu'à des époques très anciennes. Les mesures sont ensuite calibrées dans un référentiel pour obtenir une datation absolue. La racémisation des acides aminés permet de remonter entre 200000 ans (~20°C, pH neutre) et 10 millions d’années (~-10°C, pH neutre) sinon davantage si l'échantillon le permet. Cette technique de datation est utilisée depuis des dizaines d'années et fut notamment utilisée pour dater des fossiles de l'Homo rhodesiensis en 1974. Précisons que si cette méthode ne peut pas s'appliquer (c'est parfois le cas en paléontologie et en stratigraphie mais également en médecine légale), on peut également étudier la racémisation de l'acide aspartique ou de l'acide glutamique accumulé dans les protéines humaines (par ex. dans les protéines des os) dans le même but comme l'explique cet article scientifique. La technique de racémisation des acides aminés ainsi que des acides aspartique et glutamique sont exploitées en paléontologie, en stratigraphie, en paléobiologie, en paléoclimatologie, en archéologie, en taphonomie (l'étude de la fossilisation) et en néotectonique notamment afin de répondre à des questions relatives à la chronologie, la fréquence ou au taux d'évolution des processus biologiques ou de surface. En complément, l'étude de la sensibilité des acides aminés fossilisés à la température permet d'affiner les méthodes de géochronologie et de paléothermométrie. La typologie La typologie est un système de classification intuitif utilisé depuis des sièces par les chercheurs pour comparer des objets témoins entre eux de façon à les classer selon leur ressemblance afin de les rattacher à un contexte ou à une période donnée. Concrètement, si un objet inconnu ressemble à la forme et au style d'un autre objet connu de même nature, on peut supposer qu'ils datent de la même époque. Il s'agit donc d'une méthode de datation relative mais elle est fréquemment utilisée lorsqu'il est impossible de recourir à des méthodes de datation absolue ou pour identifier rapidement si le site correspond au gisement convoité (site fossilifère, archéologie sous-marine, etc). Elle permet généralement de connaître la période culturelle ou géologique à laquelle appartiennent un site ou un artefact, sans préciser la date d'occupation. On utilise cette méthode pour classifier des artefacts tels des outils lithiques, des poteries, des céramiques et des objets métallurgiques, car il est souvent possible de comparer les matériaux, leur morphologie, les techniques de décoration et les motifs entre les différents artefacts. La typologie ne s'applique pas à la découverte d'un objet unique puisqu'il n'existe pas de base de comparaison. On pourrait juste tenter de le classifier par dissemblance.
Dans le même contexte, la typochronologie permet de dater par comparaison des objets de même type, même si les méthodes artisanales, les habitudes culturelles ou les modes ont évolué dans le temps. Cette méthode reste toutefois peu précise et sujette à interprétation car la même industrie peut aussi se développer à d'autres époques selon la région. Ainsi les pointes de flèches à encoches découvertes en Californie fabriquées il y a 10000 ans (et jusqu'au XVIe siècle) peuvent être 6000 ans plus anciennes que celles découvertes en Europe. En revanche, la découverte en France d'un silex taillé en feuille de laurier de type Volgu est tellement typique que tous les archéologues peuvent affirmer qu'elle date de l'époque solutréenne (20000-18000 ans BP). La biochronologie La biochronologie combine certaines branches de la biologie et spécialités de la géologie avec la typologie pour affiner la datation. Elle permet de comparer les différents échantillons organiques aux fossiles de faune et de flore présents dans la même strate et d'évaluer s'ils ont éventuellement vécu à la même époque. Elle peut donc pratiquement faire appel à des chercheurs spécialisés dans n'importe quelle branche de la paléontologie : paléobiologie, paléobotanique, paléogénétique, archéobiologie, sédimentologie, etc. Ainsi, pour préciser l’âge de Toumaï, les scientifiques se sont basés sur les restes fossiles de Proboscidiens et de Suiformes trouvés dans les mêmes conditions, sur le même site. Voyons à présent quelles sont les disciplines et spécialités qui viennent compléter le travail des paléontologues en apportant des informations sur les paléo-écosystèmes, le paléoclimat et l'impact éventuel de l'homme sur l'environnement. Deuxième partie
| ||||||||||||||||||||||||||||||||||||||||










{kind=link}
{kind=link}
{kind=link}