|
|
|
Bactéries et virus
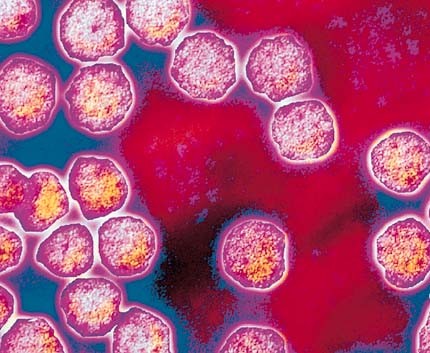
Les virus (III) Les virus[2] n'ont été découverts qu'au XXe siècle, et plus exactement dans les années 1930, grâce à l'invention du microscope électronique. En effet, la plupart des virus sont invisibles dans un microscope optique; ils sont mille fois plus petits que les bactéries avec une taille moyenne de l’ordre de 100 nm soit 0.0001 mm, c'est 1000 fois plus petit qu'un cheveux (30-100 microns). Le plus petit virus (sa capside) mesure 17 nm (circovirus porcin de type 1) et le plus grand virus mesure 440 nm (Megavirus chilensis). Ce type de microbe dispose d'une paroi de nature protéinique mais n'abrite qu'un seul acide nucléique. On y reviendra. Plus de 5000 espèces de virus ont été décrites dont plus de 2300 sont reprises dans la base ICTV. Combien y a-t-il de virus et d'espèce virales ? Les virus sont 10000 à 100000 fois moins nombreux dans notre corps que les microbes mais leurs effets peuvent être bien plus conséquents. En excluant les attaques virales qui multiplient les virus par milliards, on estime qu'un être humain en bonne santé porte plus de 300 millions de virus, essentiellement des bactériophages (voir plus bas). On les trouve principalement dans les muqueuses : les fosses nasales, dans la bouche et dans les intestins, où ils se complaisent et assurent en force notre défense contre toutes sortes de parasites. Bien sûr, certaines espèces sont pathogènes. En 1993, l'épidémiologiste Stephen Morse du Centre Médical de l'Université de Columbia estima qu'il y avait environ un million d'espèces virales de vertébrés (calculées en supposant qu'il existe environ 20 espèces virales différentes dans chacun des 50000 vertébrés de la planète). Une nouvelle étude publiée en 2013 suggère en extrapolant qu'il existe au moins 320000 espèces virales différente infectant les mammifères. D'autres extrapolations à partir du nombre de virus inconnus découverts chez les petits mammifères (rats, chauves-souris, singes, etc.) et la volaille, suggèrent qu'il existe entre 300000 et 500000 espèces de virus infectant les mammifères dont la plupart restent à découvrir. Il faut y ajouter les millions de virus qui infectent les autres organismes, y compris les plantes et les centaines de millions d'espèces de virus qui peuplent les océans. Au total, il est probable que la Terre abrite jusqu'à un milliard d'espèces de virus ! On y reviendra. Rien qu'en se limitant aux virus des mammifères, les plus à même à sauter de l'animal à l'être humain, dans l'esprit d'un virologue-enquêteur, cela représente quelque 500000 virus dont le mode d'infection et la dangerosité sont inconnus. Quand on connaît l'effet d'Ebola ou du VIH (HIV en anglais) porteur du SIDA, cela donne des sueurs froides... Mais gardons notre sang-froid et étudions littéralement à la loupe ces créatures au métabolisme très étonnant et aux effets parfois aussi spectaculaires et parfois foudroyants. D'où proviennent les virus ? Au cours du séquençage du génome de différents virus, il est apparu que certains d'entre eux disposent de gènes empruntés aux animaux. Ainsi, dans une étude publiée en 2016, des chercheurs de l'Université Vanderbilt ont annoncé la découverte d'un phage (un virus qui tue les bactéries) qui s'attaque au Wolbachia, un parasite bactérien, dont un tiers des gènes correspondent à l'ADN d'une toxine qu'on retrouve dans le venin de l'araigné veuve noire. Ceci implique qu'à l'origine ce virus a recruté ce gène chez cet animal pour faciliter sa vie : infecter les bactéries qui vivent dans les araignées et les insectes ! D'un autre côté, si les virus et autres phages sont des parasites et souvent des tueurs, la découverte de virus dans nos gènes de rebut, dans le placenta des mammifères ainsi que des macrovirus laissent à penser que tous les virus ne sont pas dangereux; certains participeraient activement au développement de la vie. Cette découverte[3] bouleversant nos théories sur l'évolution, nous y reviendrons en détail à propos du rôle des virus dans l'évolution. Définition Comme son nom latin l’indique, les virus sont de véritables poisons. Ce terme fut utilisé pour la première fois en 1890 pour qualifier des agents infectieux qui étaient plus petits que les bactéries. D'un point de vue purement taxonomique, on définit les virus comme "des agents infectieux constitués par un acide nucléique ARN ou ADN enrobé par une enveloppe protéique." En 1962, pendant qu'ils classifiaient les différents types de parasites selon leurs propriétés et leurs formes, les biologistes Donald L. Caspar et Aaron Klug ont défini les "virus" comme étant "des parasites intracellulaires obligés qui se multiplient en reprogrammant la cellule infectée pour assurer la production des composants viraux."
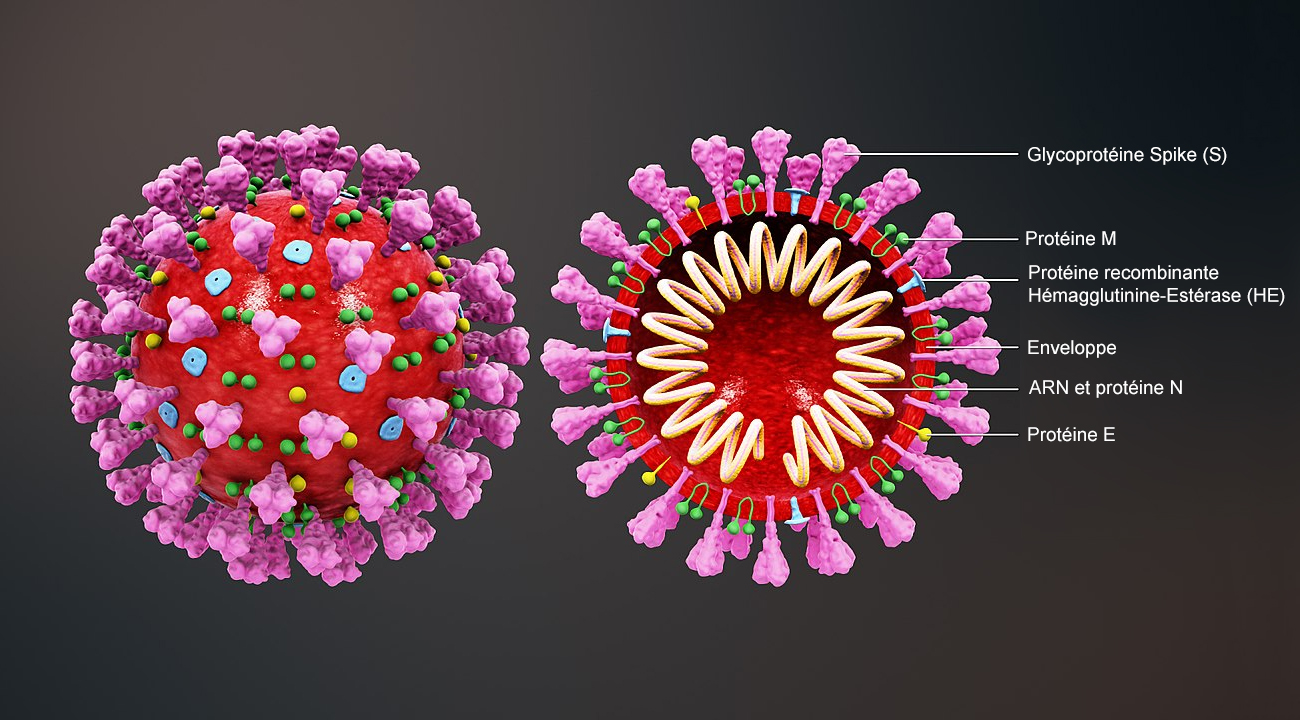
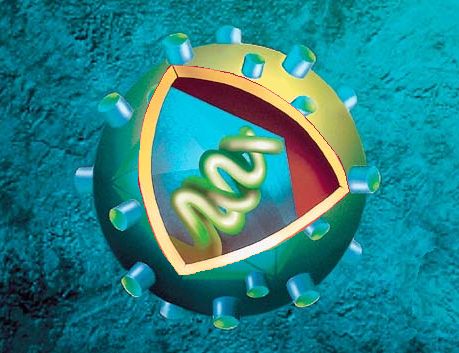
Ces descriptions impliquent que les virus sont des structures très simples - c'est relatif - composées d'un acide nucléique et d'une enveloppe destinée à protéger cette information génétique. Ajoutons que ces parasites sont incapables de se reproduire seuls et ont besoin de la machinerie cellulaire pour y parvenir. On ne peut donc pas les qualifier d'organismes vivants bien que la différence soit subtile. Pour éviter d'utiliser le terme "vivant " on peut aussi parler de "charge active". Dans ce contexte "virus" est un terme générique vernaculaire comme "humain" regroupe les hommes et les femmes, tout en sachant le genre "virus" comprend de nombreuses groupes et espèces. Virus ou virion ? D'un point de vue microbiologique, on parle de "virus" ou de charge active lorsque l'agent infectieux se trouve à l'intérieur d'une cellule hôte et est complet (non fragmenté), c'est-à-dire constitué de son matériel génétique (ARN ou ADN), de sa capsule protéique, de ses spicules protéiques et de son enveloppe lipidique. Dès qu'il émerge ou bourgeonne de la cellule à la fin de son cycle de vie (voir plus bas) ou lorsqu'il se trouve à l'extérieur de la cellule, on le qualifie de "virion" ou de particule ou charge inerte. Le virion ne contient pas de cytoplasme ni de ribosomes. Structure cellulaire Les virus n'ont pas de noyau contenant le matériel génétique, donc ils sont incapables de se reproduire par mitose (division cellulaire). Ils ne disposent pas non plus d'enzymes qui jouent le rôle de catalyseur pour produire l'énergie nécessaire à leur métabolisme. Ils ne possèdent pas de chromosomes porteur de l'information génétique (les gènes contenant les nucléotides). En revanche ils renferment un acide nucléique. Dans ce cas comment se développent-ils ? C’est le biochimiste américain Wendell Meredith Stanley qui découvrit en 1935 que cet acide nucléique était formé d’un ARN chez les rétrovirus et d’un ADN chez les virus de nature ADN. Ce génome contient généralement une dizaine de gènes. Le record est détenu par un Pandoravirus décrit en 2013 contenant entre 1900 et 2500 gènes, soit 10% du génome humain ! Ce génome viral est généralement protégé par une capside faite de protéines qui permet de différencier les virus. La capside est parfois enveloppée dans une membrane plasmique appelée péplos (cas du SARS-CoV-2, des Influenzavirus de la grippe, du papillomavirus de l'hépatite, du VIH, etc.) hérissée de spicules produisant des glycoprotéines. Ces spicules comme la protéine S du SARS-CoV-2 permet au virus de se lier à un récepteur spécifique de la cellule hôte, dans ce cas ci ACE2 afin d'infecter la cellule.
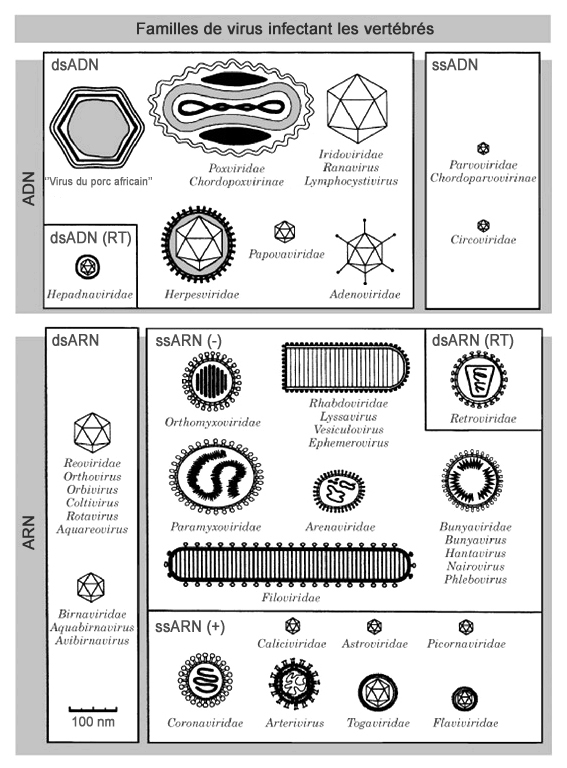
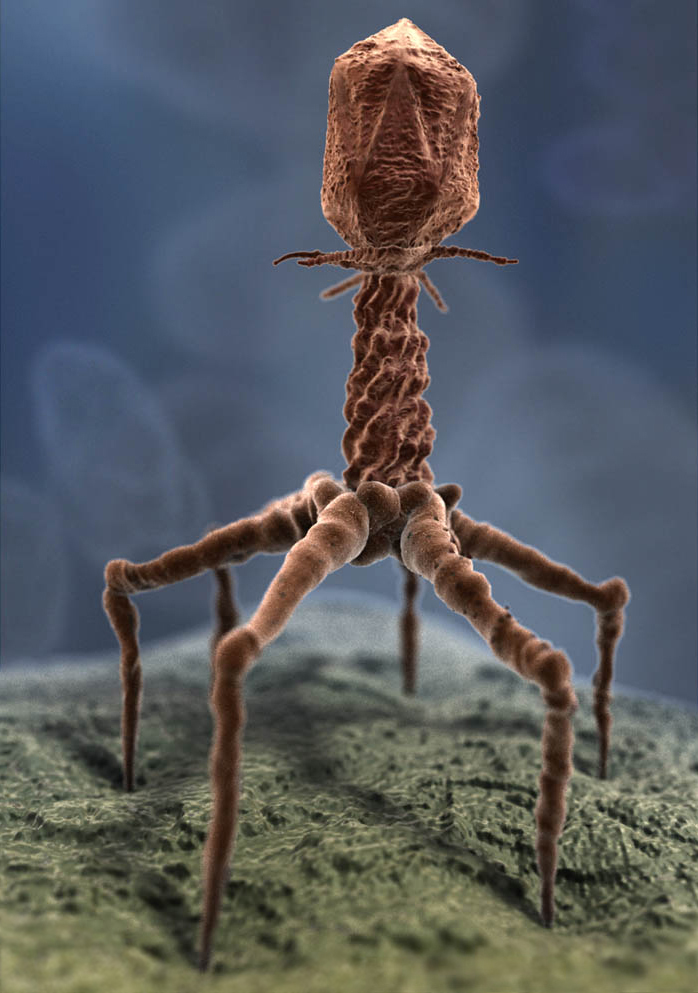
Propriétés générales des virus Dire qu'un virus est vivant est un abus de langage mais le sujet fait toujours débat (cf. cette thèse). Il faut en fait comprendre que cela représente la période (la demi-vie) durant laquelle la charge virale reste active. Tous les virus parasitent les cellules, qu'elles soient végétales ou animales car ils ne peuvent se développer et se reproduire en dehors de celles-ci. En effet, tout le matériel génétique qu’ils renferment est inactif et ils ont besoin des fonctions enzymatiques et métaboliques d’une cellule pour exécuter leur programme génétique. Les virus présentant une structure interne hélicoïdale tel le phage T4 peuvent atteindre un diamètre de 0.1 mm (la taille d'une bactérie) et sont bactériophages, c’est-à-dire qu’ils sont à ce point pervers qu’ils se développent au sein même des bactéries et les détruisent. Leur action à tous les aspects d'une extermination globale : tous les 2 jours, 50% des bactéries présentes sur terre meurent sous l'assaut des bactériophages ! La méthode d'infection d'un virus est simple et redoutablement efficace : le virus détourne à son profit les fonctions du métabolisme cellulaire. Grâce à des antigènes de surface (H et N, voir plus bas), le virus de la grippe par exemple va se lier aux récepteurs présents sur la paroi d’une cellule saine, pénétrer sa membrane et injecter son ARN dans la cellule hôte. L'ARN va se combiner avec l'ADN cellulaire pour être transcrit sous la forme de nouveaux virus qui iront infecter de nouvelles cellules. On y reviendra. Mais d’autres scénarii existent. Certains virus pénètrent dans la membrane cellulaire mais ne tuent pas leur hôte. D’autres sont “silencieux” et infectent la cellule sans créer apparemment de dommage cellulaire. Statistiquement, 50% des agents pathogènes présents chez l'homme proviennent des animaux (singe, rat, chauve-souris, etc), la contamination se faisant par contact physique (avec la salive, le sang, l'urine, les selles). La propagation n'est pas réciproque :; si vous avez attrapé un virus, vous ne contaminerez pas votre chien ou votre chat. Tous les modes de transmission sont possibles, bien que la plupart des maladies virales se transmettent par voie orale ou fécale et par les piqûres d’insectes qui furent eux-mêmes contaminés en suçant par exemple le sang d'un animal infecté et porteur sain. Les rétrovirus, découverts en 1910 mais considérés comme des chimères pendant 60 ans[4] sont plus malins encore et se passent des ARN messagers (ARNm). Ils assurent eux-mêmes l'assemblage des protéines. Non seulement ils leurrent les cellules hôtes au moment de la reproduction mais ils les détruisent en provoquant de nombreuses maladies infectieuses à l'origine des cancers. Certains enfin, sont utilisés par les biologistes. Plus inoffensifs, ils sont toutefois stérilisés pour éviter toute félonie : ce sont les virus blancs dont le rôle, à l’instar des virus synthétiques comme les liposomes utilisés dans l’industrie cosmétique, se rallie à la thérapie génique. Classification des virus Si la taxonomie permet aux biologistes de classer les virus comme tous les organismes dans l'arbre phylogénique en taxons rangés par ordre, famille, sous-famille, genre, et espèce, les virologues ont préféré les classer de manière plus pratiques selon des caractéristiques génétiques, morphologiques et infectieuses. En général, la classification des virus concerne la dénomination des virus et l'assemblage de ces noms selon un critère donné. Dans la classification des organismes biologiques, ce critère est souvent basé sur des relations phylogénétiques (évolutives). Cependant, il n'y a aucune preuve que les virus possèdent un ancêtre commun ou sont de quelque manière phylogénétiquement liés. Néanmoins, la classification selon le système de Linné (1735-1758) en familles, genres et espèces a été partiellement couronnée de succès. De plus, sur la base des hôtes que les virus infectent, la première grande division utilisée dans les schémas de classification des virus implique la séparation des virus en virus de vertébrés, virus d'invertébrés, virus végétaux, virus bactériens et virus d'algues, de champignons, de levures et de protozoaires (bien que les familles de virus peuvent appartenir à plusieurs de ces classes). Au sein de ces classes, d'autres critères de subdivision sont utilisés, notamment la morphologie générale (par exemple, la présence ou l'absence d'enveloppe), la nature du génome (ADN ou ARN), la structure du génome (simple brin (ss) ou double brin (ds), linéaire ou circulaire, fragmenté ou non fragmenté), les mécanismes d'expression des gènes et de réplication virale (ARN de sens positif ou négatif), les relations sérologiques, la sensibilité de l'hôte et des tissus et les pathologies qu'ils provoquent (symptômes ou type de maladie). Le sujet étant encyclopédique, pour plus d'informations sur la classification des virus lire notamment la classification ICTV ainsi que les article du Comité exécutif de l'ICTV (2020), M.M.Pierce (2020) et Elliot J Lefkowitz et al. (2017) parmi d'autres. 1. Classification de Baltimore En 1971, le virologue David Baltimore proposa dans un article de 7 pages intitulé "Expression of ANimal Virus Genomes", une première classification des virus selon la nature de leur acide nucléique et leur expression, c'est la classification de Baltimore. Les virus à double brin d'acide nucléique (double stranded ou ds) sont appelés bicaténaires, les virus à simple brin d'acide nucléique (single stranded ou ss) sont appelés monocaténaires. Les virus sont classés en 7 groupes reprenant plusieurs familles de virus :
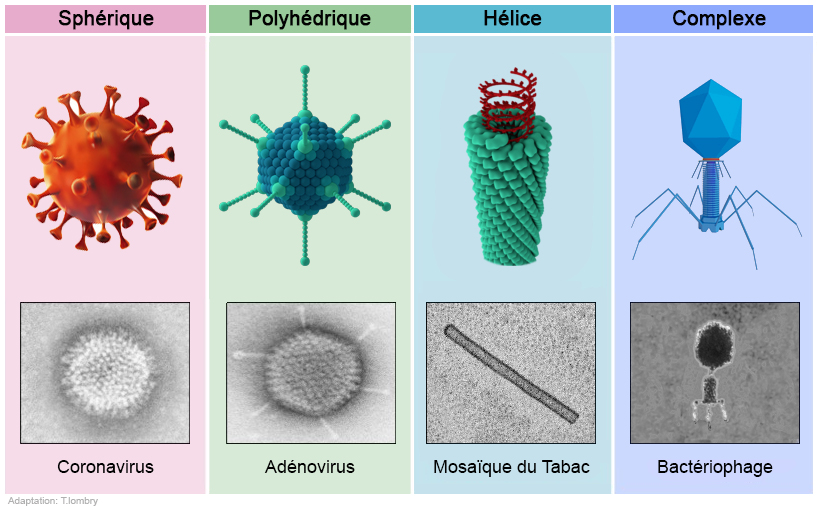
2. Classification morphologique Les virus sont également classés sur base morphologique : - La symétrie de l'enveloppe extérieure (la capside protéique) - la présence ou l'absence de membrane lipidique (péplos) - Les dimensions du virion et de la capside. Le tableau suivant résume les caractéristiques des virus.
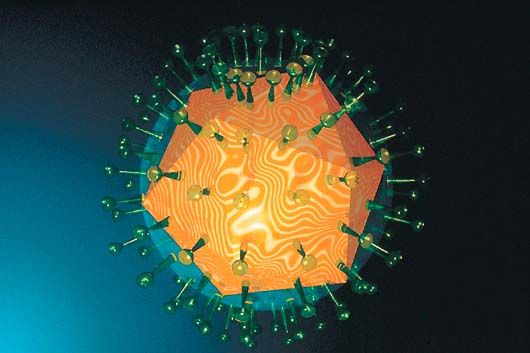
3. Description des antigènes H et N des virus grippaux (Influenzavirus) Les virus pathogènes, des coronavirus (et les 180 autres) à l'origine des pneumonies virales ou de la bronchite au genre Ebola en passant par le VIH se distinguent également par leurs antigènes, ces substances étrangères à l'organisme hôte qui vont déclencher une réaction du système immunitaire afin de les éliminer. Dans le cas des virus de la grippe par exemple (au pluriel car il existe de nombreuses souches), le genre Influenzavirus, de la famille des orthomyxoviridae, comme on le voit ci-dessous à gauche, le brin du génome d'ARN est divisé en 8 segments sur lesquels sont codés 11 protéines (comprennent une polymérase assurant la synthèse de l'ARN et des protéines de structure ou d’enveloppe). L'unicité de ces virus est déterminée par deux marqueurs, des protéines d'antigènes présentes à leur surface : l'hémagglutinine (H ou HA) et la neuraminidase (N ou NA) qui déterminent leur pouvoir infectieux. A ce jour, il existe 19 sous-types H et 9 sous-types N qui peuvent se combiner, donnant un total de 171 combinaisons virales potentielles. Seule une poignée d'entre eux a infecté l'être humain. Concrètement, les virus de la grippe tels celui illustré ci-dessous à gauche présente à sa surface de petites spicules H et N. La dénomination antigénique A(H1N1) par exemple de la grippe saisonnière fait référence à la souche d'Influenza de type A la moins contagieuse (mais qui peut tout de même tuer des millions de personnes). En revanche, la souche H5N1 comme le virus Ebola, est très contagieuse et tue un homme en quelques semaines si aucun traitement n'est prodigué ! Enfin, le sous-type H7N7 par exemple est hautement pathogène (HP) mais comme H1N7, jusqu'à présent il reste confiné chez les animaux (oiseaux, procs, chevaux, etc) ce qui ne veut pas dire qu'il ne peut pas infecter l'homme. On reviendra sur les modes de transmission. A voir : Molecular Art, David S. Goodsell
Le pouvoir pathogène d'un virus est relatif car même s'il représente un agent pathogène mortel pour une espèce il ne l'est pas obligatoirement pour une autre. Non seulement la souche de ce virus doit être transmissible d'une espèces à l'autre mais elle doit aussi vaincre le système immunitaire de son hôte. Généralement les antigènes viraux de surface sont reconnus par les agents du système immunitaire de l'organisme infecté qui vont déclencher une réaction de défense et l'éliminer. Si le virus veut conquérir son nouvel hôte il doit donc soit compter sur une baisse des défenses immunitaires de son hôte soit subir une mutation. Nous allons y revenir. 4. Virus enveloppé et virus nu Les influenzavirus ou les coronavirus sont des virus enveloppés, c'est-à-dire que la capside protéinique est entourée d'une structure souple, le péplos, une membrane dérivée des membranes cellulaires, cytoplasmique ou nucléaire selon les virus. Cette enveloppe étant fragile, un virus enveloppé comme le SARS-CoV-2 peut plus facilement perdre son pouvoir infectieux dans le milieu extérieur ou dans le tube digestif par exemple que les virus nus qui résistent beaucoup plus longtemps. 5. Voies de transmission Généralement, les virus ne franchissent pas les barrières interspécifiques, ce qu'on appelle la zoonose, par exemple des moustiques à l'homme, des chauves-souris à l'homme ou du chien à l'homme. En revanche, nous savons que les animaux sont des réservoirs naturels de virus et autres pathogènes. On estime que l'ensemble des animaux (les humains exclus) abrite 1.5 million d'espèces virales inconnues et potentiellement transmissibles à l'être humain ! Ces animaux sont des vecteurs dits passifs car ils peuvent conserver et amplifier une charge virale. En effet, un moustique par exemple peut piquer et aspirer le sang d'un animal contaminé par des flavivirus, les conserver dans son sang sans être infecté puis les injecter dans un autre animal lors d'une piqûre. Cet animal peut-être un humain qui dans ce cas ci contractera la fièvre jaune, la dengue (toutes deux transmises par les moustiques du genre Aedes) ou la fièvre West Nile (transmis par les moustiques du genre Culex). Notons que la paludisme n'est pas transmis par un virus mais par un parasite du genre Plasmodium via le moustique du genre Anophèle. Si une contamination par zoonose se produit, il s'agit toujours de conditions très particulières. Nous savons par exemple que le rétrovirus VIH du SIDA a été transmis du singe à l'homme par contact. Pour être transmissible, la cellule hôte doit disposer du récepteur particulier afin de permettre au virus de pénétrer dans la cellule. Sans cette "serrure" adaptée à cette "clé virale", le virus ne peut pas attaquer son hôte et risque à tout moment d'être détruit par les défenses de notre système immunitaire.
Lors des épidémies revient souvent la question de la transmission d'une maladie à son chien ou son chat. C'est très peu probable. En effet, les virus humains et ceux attaquant nos animaux domestiques sont différents. On parle bien du même virus, par exemple l'herpès ou le coronavirus, mais il existe plusieurs familles ou groupes : alpha, bêta, gamma, delta, etc comme le montra l'équipe de P.C. Woo en 2012. Le virus HHV-1 de l'herpès buccal est de type alpha. Malheureusement, c'est le même qu'on retrouve chez le singe. Il peut donc le transmettre à l'homme par contact notamment. Plusieurs coronavirus humains mais aussi de chauves-souris sont du genre alpha. En revanche, le coronavirus des canins et félins est du genre alpha et n'est pas transmissible à l'homme et vice-versa. Inversement, le Covid-19 sur lequel nous reviendrons est un bêtacoronavirus. Autrement dit, nous ne pouvons pas contaminer nos chiens et chats car leurs cellules ne sont pas munies des récepteurs spécifiques. Si en théorie, ils sont donc insensibles à ce pathogène, on connaît une poignée de cas où des animaux furent contaminés par l'homme. En général, ils s'auto-immunisent ou présentent des symptômes bénins faciles à soigner. Mais cela reste anecdodique. On reviendra sur les zoonoses page suivante. Quant aux voies de transmission proprement dites, comme on le voit dans le tableau ci-dessous, les virus ont développé de nombreuses stratégies pour atteindre leur but. Mais généralement, ces stratégies sont exclusives ou en tous cas elles sont optimisées pour un certain mode de transmission. Ainsi, en théorie les coronavirus comme le Covid-19 peuvent se transmettre par voie aérienne mais en pratique, étant enveloppé, leur péplos ne résiste pas longtemps dans l'air (quelques dizaines de secondes à quelques heures) et leur charge virale est rapidement détruite.
Ceci dit, les maladies qui se transmettent le plus rapidement et le plus facilement sont celles dont les virus se propagent dans l'air, comme les Influenzavirus (grippe) ou les coronavirus (rhume, SARS, MERS, SARS-CoV-2, etc). Un contact physique avec une personne contaminée par un tel virus n'est pas nécessaire pour le transmettre. Chacun sait qu'on peut attraper un rhume ou la grippe en inhalant des particules virales projetées par une personne contaminée, même sans la toucher (cf. les voies de transmission du Covid-19). En revanche, les maladies transmises par les fluides corporels, comme Ebola ou le VIH, ne sont pas aussi faciles à contracter ou à transmettre. En effet, il faut entrer en contact avec du sang, de la salive ou d'autres fluides corporels infectés pour le contracter. Rappelons que le SIDA n'est pas une maladie contagieuse comme peut l'être la grippe saisonnière ou la Covid-19. Le VIH ne peut pas se transmettre par un baiser, un éternuement, une toux, une piqûre d'insecte, le partage d'un verre, d'un vêtement, etc. Les maladies aéroportées ont généralement une contagiosité "Ro" ou taux de reproduction de base (cf. la modélisation des épidémies) plus élevé que celles transmises par contact direct et se transmettent plus facilement par contact. 6. Caractéristiques complémentaires En 1980, l'OMS proposa d'ajouter six caractéristiques complémentaires à la description des virus : - le type d'antigène (par ex. A, M, N, S) qui distingue la nature des protéines que portent le virus - l'hôte d'origine (l'animal s'il est différent de l'homme) - l'origine géographique (par ex.Taiwan, etc) - le nombre de souches ou "strains" (par ex. 15, 7, etc) - l'année d'isolation (par ex. 57, 2009, etc) - pour les virus Influenza A, la description des antigènes H et N (par ex. (H1N1), etc). Cela donne par exemple les descriptions suivantes : 1. A/duck/Alberta/35/76 (H1N1) désigne un virus isolé chez le canard 2. A/Perth/16/2009 (H3N2) désigne un virus Influenza d'origine humaine. Chez la plupart des organismes la molécule d'ADN se compose de 4 désoxyribonucléotides ou bases nucléiques qui sont presque toujours l'adénine (A), la guanine (G), la thymine (T) et la cytosine (C). La molécule d'ARN utilise presque le même alphabet, mais utilise l'uracile (U) à la place de la thymine. En 1977, un groupe de scientifiques découvrit en Russie le premier cyanophage, c'est-à-dire un virus qui s'attaque aux cyanobactéries, dont l'adénine (A) avait été remplacée par la 2-aminoadénine (Z) également appelée 2,6-diaminopurine. Autrement dit, l'alphabet génétique qui se compose généralement des lettres A, G, T et C chez la plupart des organismes es composé des lettres Z, G, T et C chez ces virus à ADN cyanophages.
Pendant des décennies, il s'agissait de la curiosité la plus étrange de la biologie et on savait peu de chose de l'effet de la substitution de cette nucléotide sur le virus. À la fin des années 1980, les chercheurs découvrirent que ce nucléotide Z donnait en fait certains avantages au virus : il était plus stable à des températures plus élevées, permettant au brin d'ADN de se lier plus précisément au deuxième brin d'ADN après réplication. De plus, le "génome Z" résiste mieux aux protéines de défense de la bactérie qui normalement détruisent l'ADN viral. Il permet donc à ces bactériophages d'échapper aux bactéries. Dans trois études distinctes publiées dans la revue "Science" en 2021 (cf. S.Zhao et al., 2021; P.Kaminski et al., 2021 et P.Marlière et al., 2021) et dans une quatrième publiée dans la revue "Nature" en 2021 par Pierre Kaminski et de l'institut Pasteur et ses collègues, les chercheurs ont découvert comment le nucléotide Z se retrouve dans le génome de ces cyanophages et utilise la machinerie cellulaire pour se répliquer. Le génome Z est très stable. Le nucléotide Z se différencie de l'adénine par la présence d'un groupe amine (NH2) supplémentaire sur l'un des atomes de carbone. Lorsque les deux brins d'ADN se réunissent pour former la double hélice inversée, deux liaisons hydrogène lient les bases A à T et trois liaisons hydrogène lient C à G. Mais lorsqu'on substitue A par Z, en raison de la présence de cette amine, la molécule s'apparie avec la thymine ou avec l'uracile en formant trois liaisons hydrogène, renforçant ainsi le lien. A ce jour, c'est la seule chaîne d'ADN qui modifie la liaison hydrogène. Selon les chercheurs, il n'est pas surprenant que le génome Z soit peu répandu chez les espèces actuelles. Le génome Z crée un ADN très stable, mais peu flexible. En effet, dans de nombreux évènements biologiques, tels que la réplication de l'ADN, il faut décompresser et séparer le double brin d'ADN et la liaison hydrogène supplémentaire rend la décompression plus difficile. Selon Zhao, "Je pense qu'il est plus adapté aux environnements chauds et difficiles, mais pas à cet environnement plus confortable que nous connaissons actuellement." Selon les chercheurs, il est possible que ce génome Z ait existé très tôt dans l'histoire de la Terre. Il aurait pu conférer un avantage aux premières formes de vie, leur permettant de se développer dans l'environnement hostile de la Terre primitive. La stabilité du génome Z en fait un candidat idéal pour certaines technologies. Maintenant que les chercheurs savent quelles protéines utilise le virus pour fabriquer ces génomes Z, les scientifiques peuvent les fabriquer eux-mêmes à grande échelle. Les projets ne manquent pas. Selon les chercheurs, le génome Z permettrait d'améliorer la thérapie phagique, une méthode de traitement des infections bactériennes qui utilise des bactériophages, généralement lorsque les bactéries développent une résistance aux antibiotiques. Il pourrait aussi être utilisé pour améliorer la longévité et la capacité de ciblage des brins d'ADN utilisés en thérapie génique. Enfin, les chercheurs pourraient étudier les avantages que procure le génome Z si on l'incorporait dans les cellules. Mais en attendant, il reste beaucoup de questions en suspens. Zhao et ses collègues vont étudier la structure tridimensionnelle du génome Z et vérifier s'il présente des différences par rapport à celle de l'ADN normal, tandis que l'équipe de Kaminski va explorer les autres avantages que ce génome Z procure aux cyanophages. On ne sait pas non plus si le génome Z peut également former des brins d'ARN ni s'il peut s'intégrer dans les gènes de l'hôte bactérien. Ce qui ressort clairement de ces études, c'est que le génome Z est plus répandu que nous ne le pensions et présente probablement une histoire évolutive très intéressante à étudier. Enfin, on a découvert dans des météorites carbonées des molécules de 2-aminoadénine et d'autres acides aminés, suggérant que des bases nucléiques peuvent se former dans les astéroïdes notamment, ce qui est une très bonne nouvelle pour les exobiologistes (cf. M.P. Callahan et al. (en PDF), 2011; A.S. Burton et al. (en PDF), 2012). On y reviendra en bioastronomie. De nouveaux virus à ARN dans les océans Entre 2009 et 2013, une équipe internationale de chercheurs dirigée par l'Université d'État de l'Ohio en collaboration avec des chercheurs de l'EMBL, du CNRS, du CEA, de l'ENS-PSL et de la Fondation Tara Ocean, un projet international visant à étudier l'impact du changement climatique sur les océans, parcourut plus de 75500 miles marins soit 140000 km sur tous les océans du monde et préleva 35000 échantillons d'eau et effectua ~13000 mesures contextuelles dans 121 écosystèmes marins différents et à trois profondeurs d'eau. Les chercheurs ont ensuite analysé les séquences génétiques du plancton, qui sont des hôtes habituels des virus à ARN, recherchant spécifiquement un gène ancestral appelé RdRp, qui se trouve dans tous les virus à ARN (y compris le SARS-CoV-2 et recherché lors des tests RT-PCR), mais est absent des autres virus et cellules. Ils ont identifié plus de 44000 séquences possédant ce gène qui remonte à plusieurs milliards d'années et qui a subi de nombreuses mutations. Vu l'ampleur du travail d'analyse, les chercheurs ont fait appel à l'I.A. et spécifiquement à l'apprentissage automatique, grâce auquel ils ont identifié 5504 nouvelles espèces de virus à ARN appartenant aux cinq phyla existants, ainsi que cinq nouveaux phyla, que les chercheurs ont nommés Taraviricota, Pomiviricota, Paraxenoviricota, Wamoviricota et Arctiviricota. Les espèces de virus du phylum Taraviricota étaient particulièrement abondantes dans les eaux tempérées et tropicales, tandis que les virus du phylum Arctiviricota sont abondants dans l'océan Arctique. Les résultats de leurs analyses furent publiés dans la revue "Science" en 2022.
Selon les chercheurs, comprendre comment le gène RdRp a divergé au fil du temps pourrait conduire à une meilleure compréhension de la façon dont les premières formes de vie ont évolué sur Terre. Selon Ahmed Zayed, chercheur en microbiologie à l'Université d'État de l'Ohio et principal auteur de cet article, "RdRp est censé être l'un des gènes les plus anciens - il existait avant que l'ADN ne soit nécessaire. Nous ne retraçons donc pas seulement les origines des virus, mais également les origines de la vie." Les travaux des chercheurs ont permis de porter le nombre d'espèces virales marines de 16000 à ~21500 mais selon le biologiste Chris Bowler du CNRS, il pourrait exister jusqu'à 1 milliard d'espèces virales dans les océans ! Malheureusement, vu le coût très élevé de la recherche en virologie (l'étude d'un seul des 58 virus que porte la chauve-souris Pteropus giganteus - le renard volant d'Inde - coûta 1.2 million de dollars, cf. virology), on sait déjà qu'il sera impossible de les étudier tous, ni même de les inventorier. Seul le temps pourra éventuellement y pourvoir. De nouveaux virus découverts au Tibet Nous verrons à propos de la survie dans le permafrost que des dizaines de virus furent également découverts dans un glacier au Tibet dont 28 nouveaux genres. Les chercheurs estiment que le changement climatique pourrait notamment libérer d'anciens virus pathogènes des glaciers. Des obélisques dans le système digestif Un nouveau type d'entité virale appelé "obélisque" résidant dans les bactéries fut découvert en 2024 dans le système digestif humain dont on ignore s'il affecte ou non la santé et les gènes humains (cf. I.N. Zheludev et al., 2024; Nature, 2024). On y reviendra à propos du rôle des virus dans l'évolution. Cycle de vie des virus Les virus peuvent disparaître, la souche ou le variant s'éteint, mais ils peuvent également muter et diverger (changement dans les nucléotides) très facilement et réapparaître de manière saisonnière durant des décennies sinon davantage. Par quel stratagème agissent-ils ainsi ? Les virus à ARN qui comptent aussi parmi les plus dangereux (Influenza, VIH, Ebola, SARS-CoV-2, etc) sont généralement constitués d'un seul brin d'ARN qui ne leur permet pas de disposer des systèmes de détection et de correction d'erreurs comme les virus à ADN qui contiennent des polymérases. Ces dernières sont des enzymes qui se servent de la complémentarité des bases pour synthétiser un nouveau brin et vérifier en même temps leur exactitude. En leur absence, les virus vont se reproduire pour transmettre leur message empoisonné mais soit la copie sera viable soit le virus mourra.
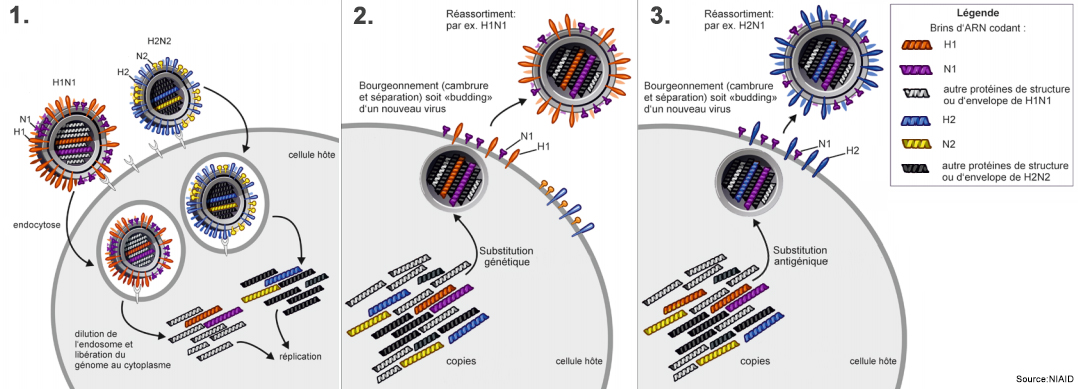
Seuls les coronavirus comme le SARS-CoV-2 disposent d'un système de correction d'erreurs grâce à l'exonucléase, une enzyme qui détruit les malfaçons dans les nucléotides virales. Elle est même capable d'identifier les molécules antivirales de médicaments comme la Ribavirine, rendant ces virus plus résistants. Si la nouvelle génération ou variant d'un virus est fonctionnelle (pour ne pas dire viable), du fait des erreurs de transcription elle peut subir des mutations aléatoires. Ce phénomène rend également le virus plus résistant car en changeant d'identité, les médicaments ne reconnaissent plus le virus original, d'où la difficulté de l'éradiquer ou du moins de le rendre inoffencif. Il devient ainsi un "serial killer aux 1000 visages" que des légions de laborantins-enquêteurs doivent pister tout au long de leur vie pour éviter qu'il ne disparaisse dans la nature et y poursuive ses méfaits (cf. l'évolution du Covid-19). Si les virus (leur charge virale) n'ont pas "conscience" de se battre contre les remèdes et les vaccins, leur facilité déconcertante à résister aux médicaments en l'espace de quelques générations - concrètement en moins d'un an -, semble leur donner des facultés d'adaptation. Face à ces mutations, les vaccins en sont réduits à ne considérer que les éléments immuables ou qui changent peu dans le patrimoine génétique des virus. Cycle de vie de l'Influenzavirus Alors qu'il n'est pas vivant, comment un virus contamine-t-il son hôte ? Dans le cas des Influenzavirus ou virus grippaux, comme on le voit sur les schémas ci-dessous, les protéines antigènes de surface H et N jouent un rôle essentiel dans la propagation et la multiplication du virus. L’hémagglutinine (H) permet au virus de se fixer sur la cellule hôte en s'attachant au récepteur correspondant, l'acide sialique. Le virus perce ensuite la membrane et s'enfonce dans la cellule où il forme une vésicule à l'intérieur du cytoplasme; c'est l'endocytose. L'endosome du virus va ensuite se diluer dans la cellule hôte afin de libérer son génome qui va ainsi pouvoir tirer profit de l'usine génétique de son hôte pour fabriquer de nouveaux virus par réplication. L'ARN va se combiner avec l'ADN cellulaire pour être transcrit par l'ARN-messager sous la forme de nouveaux virus. Cette phase de copie ne dure que quelques heures.
Lorsque la réplication est terminée, pour sortir de la cellule infectée, chaque virion va se coller à la membrane cellulaire et former un bourgeonnement. Il va ensuite utiliser sa neuraminidase (N) pour casser la liaison entre le virion et la membrane, provoquant une cambrure puis sa séparation de son hôte, ce qu'on appelle la lyse ou le "budding" d'un nouveau virus, qui est à présent libéré et peut infecter d'autres cellules. La lyse détruisant la membrane plasmique protégeant la cellule du monde extérieur, en se libérant le virus tue parfois son hôte et peut donc entraîner une destruction massive de cellules saines quelquefois accompagnée d'effets secondaires ainsi que nous l'avons évoqué. Voici le cycle de vie du SARS-CoV-2 et la manière dont il se réplique dans une cellule hôte. Les mutations C'est durant la phase de réplication/transcription que le virus peut subir une mutation de certains acides aminés de son génome. Il peut s'agir d'une substitution, d'une délétion ou d'une insertion d'un ou de plusieurs acides aminés. En effet, parfois la réplication est associée à des erreurs de transcription : des segments du génome viral initial sont recombinés différemment mais les segments codants restent inchangés. En effet, les bases ou nucléotides sont prises par 3 pour être traduites en acides aminés et un changement chez l'une d'elles n'est pas toujours accompagné d'un changement de la fonction de la protéine (cf. la synthèse des protéines). Il s'agit d'une mutation silencieuse car sans effet notable. Dans ce cas, les propriétés des virions ou cellules filles sont les mêmes que celle du virus initial. La souche virale garde sa dénomination originale, par exemple H1N1. Dans certains cas, au cours de la mutation antigénique, des segments du génome viral initial sont combinés différemment lors de la réplication et forment des segments codants offrant au virion de nouvelles propriétés. Dans ce cas, la mutation est exprimée. Le virus reçoit une nouvelle désignation, par exemple le variant H1N2. Entre les deux cas, le virus peut subir des mutations redondantes codant pour les mêmes acides aminés. Il peut aussi subir des mutations délétères qui ont un effet mortel sur le virus. Le taux de mutation est très élevé chez les virus. En effet, l'ARN polymérase n'ayant pas de système de contrôle et de réparation des erreurs de réplication comme l'ADN polymérase humain, un virus a 1 chance sur 1000 à 10000 de subir une mutation; c'est 1 million de fois plus fréquent que chez les mammifères, ce qui explique la facilité avec laquelle les virus peuvent déjouer et tromper le système immunitaire. Mais le SARS-CoV-2 avec ses 29 kbases et ses 15 gènes est déjà plus perfectionné et plus stable. En général, un virus à ARN comme le SARS-Cov-2 mute 1 million de fois par cycle viral soit environ toutes les 6 heures. Mais cela ne veut pas dire que toutes les mutations changent radicalement l'expression des gènes et l'action du virus. Ainsi le variant d'un virus peut par exemple présenter 20 nouvelles mutations par rapport à la souche originale mais seules une ou deux mutations peuvent lui offrir un avantage pour devenir plus infectieux ou pour échapper au système immunitaire de son hôte. Les autres mutations ont généralement peu d'effets. C'est la raison pour laquelle un vaccin antiviral peut protéger une personne contre plusieurs variants d'un même virus. On reviendra sur les variants à propos du SARS-CoV-2. Concernant le SARS-CoV-2, il présente un taux de mutation d'environ 25 mutations par an (en juin 2021) soit 2 fois plus lent que les virus Infuenza A. Cela s'explique en partie par la taille deux fois plus longue du génome du SARS-CoV-2 par rapport à ceux de la grippe saisonnière. Pour les virologues et les biochimistes, ces différentes mutations et leurs répétitions peuvent révéler des indices importants sur la biologie du virus. Dans le cas du SARS-CoV-2, même si certains éléments des nucléotides de la protéine S ont muté, la plupart du temps elle peut continuer à remplir sa fonction, même avec la substition de plusieurs acides aminés. Autrement dit, malgré cette entropie sur certaines positions de son ARN, ces mutations aussi nombreuses soient-elles, aboutissent très rarement à un changement du phénotype et de la fonction virale. C'est ce qu'on appelle la dérive ou le glissement antigénique qu'on observe également chez les virus de la grippe saisonnière par opposition à la cassure antigénique où deux variants du même virus produisent un nouveau sous-type de virus portant les antigènes des deux variants originaux. C'est par exemple le cas des virus de la grippe. Ceci explique pourquoi le vaccin fonctionne sur une grippe sur deux selon le variant en circulation. Le fait que les virus de la grippe saisonnière mutent si rapidement est précisément la raison pour laquelle on ne peut pas élaborer de vaccins efficaces plus d'une saison. Dernier chapitre
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||






{kind=link}