|
|
|
L'origine et l'avenir de l'Homme
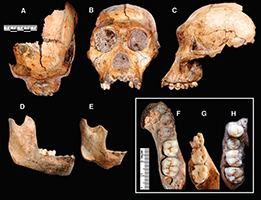
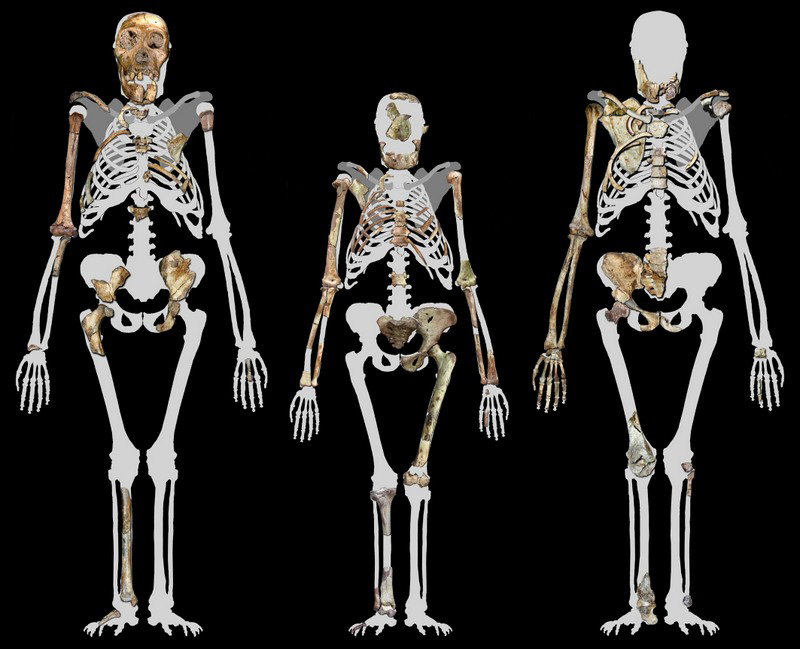
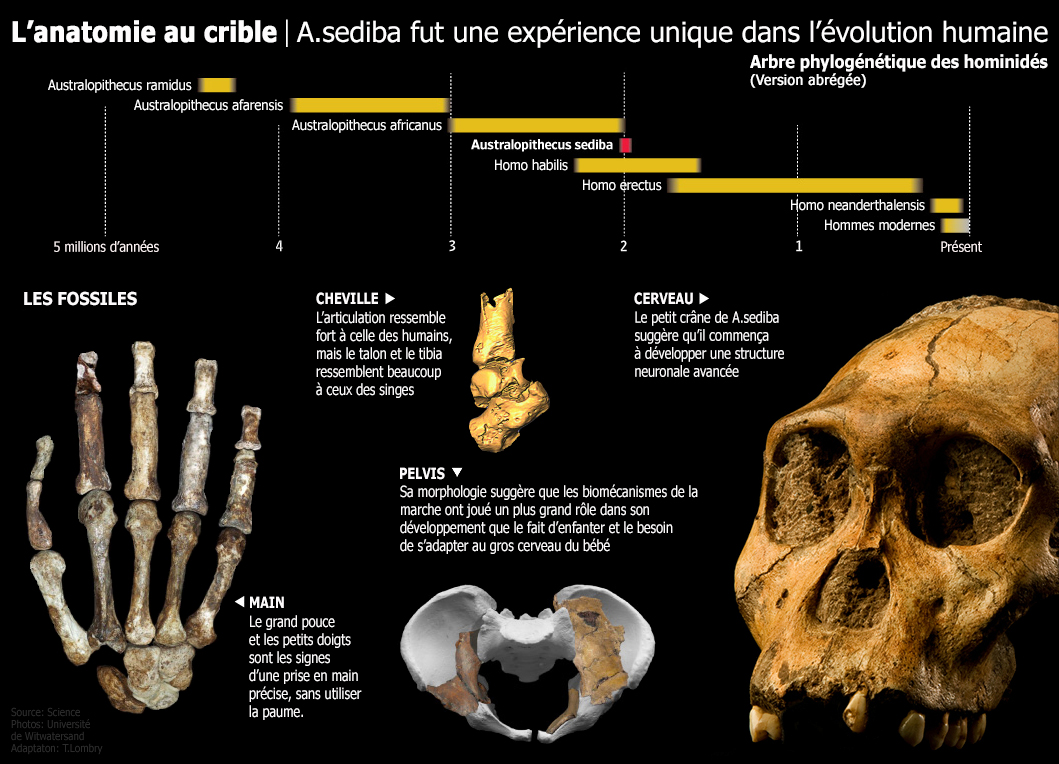
L'Australopithecus sediba : 1.98 - 1.75 million d'années (VII) Nous avons classé cette sous-espèce à part des autres Australopithèques car étant apparue plus de 2 millions plus tard que les plus anciens membre du genre, sa morphologie présente des caractéristiques très particulières qui en font un spécimen unique, hybride entre les Australopithèques et les préhumains ainsi que nous allons l'expliquer. Le premier spécimen d'Australopithecus sediba (la clavicule MH-1) fut découvert le 15 août 2008 dans la grotte de Malapa en Afrique du Sud par Matthew Berger, le fils du paléoanthropologue Lee Berger. Le squelette fut daté entre 1.980 et 1.977 million d'années. C'est donc l'un des derniers représentants des Australopithèques. Notons que ce gisement est situé à seulement 15 km des grottes de "Rising Star" où furent découvert l'Homo naledi. Les travaux d'excavations permirent également de dégager les fossiles d'autres individus dont MH-2. Au total, les fossiles de l'A.sediba représentent la collection de squelettes d'Australopithèques les plus complets que nous possédons. Cette découverte fut annoncée dans la revue "Science" en avril 2010 qui consacra par la suite tout un dossier à l'A.sediba. Actuellement nous ne possédons par d'ADN de l'Australopithecus sediba (les plus anciens sont ceux de Néandertaliens et de Dénisoviens) mais compte tenu des conditions exceptionnelles régnant dans la grotte de Malapa, Lee Berger espère un jour en découvrir. Le squelette de MH-1 comprend une clavicule droite, un crâne presque complet avec la plupart de ses dents, un bras droit, quelques côtes, quelques phalanges, des vertèbres et une partie du pelvis (bassin) et du fémur. MH-2 comprend les mêmes éléments ainsi qu'une main droite complète, une cheville et un talon, une omoplate mais le crâne est incomplet. Lee Berger estime qu'il s'agit d'une femelle. Les squelettes fossilisés d'Australopithecus sediba sont suffisamment complets pour que les scientifiques puissent observer de quelle manière les squelettes ont évolué à l'époque des premiers hommes. MH-1 est un jeune mâle encore juvénile âgé de 12 à 13 ans et mesurant 1.3 m maximum. Les Australopithecus sediba présentent un petit corps assez robuste et de longs bras. Ils ont un dimorphisme sexuel similaire à celui des humains modernes. Au début des fouilles, quand Berger examina la dentition du spécimen pour essayer d'identifier l'espèce, il constata que la mandibule était très arrondie et beaucoup moins robuste de celle de l'Homo habilis par exemple qui est plus fine et dont les molaires sont alignées. La partie frontale des molaires du spécimen était également très usée. Berger compris immédiatement que cette espèce n'est plus tout à fait un Australopithèque et se rapprochait des premiers homininés sans pour autant appartenir à ce taxon. L'Australopithecus sediba était né. Les détails de ses dents, la longueur de ses bras et de ses jambes et la partie supérieure étroite de son buste ressemblent à ceux des premiers Australopithèques mais d'autres détails de sa dentition ainsi que la largeur du bas de son torse ressemblent à ceux des humains. Ces liens indiquent que A.sediba pourrait nous apporter des informations sur les origines et l'ancêtre du genre Homo. Les modifications fonctionnelles de son pelvis indiquent des dispositions pour marcher debout bien que d'autres parties de son squelette rappellent celles d'autres Australopithèques. A.sediba n'est donc pas encore un bipède accompli.
Les mesures de la force de l'humérus et du fémur indiquent que A.sediba a une démarche plus humaine qu'un autre fossile attribué à l'Homo habilis. Ces détails suggèrent que A.sediba marchait régulièrement debout et que les modifications de son pelvis sont apparues avant les autres modifications corporelles trouvées plus tard chez les spécimens du genre Homo. Toutefois, malgré ces changements morphologiques dans le pelvis et le crâne, d'autres éléments du squelette de A.sediba montrent un corps similaire à celui des autres australopithecines présentant de longs membres supérieurs et une petite capacité crânienne. Les fossiles montrent aussi que les modifications du pelvis et de la dentition sont apparus avant les modifications de la proportion des membres ou de la capacité crânienne. Steve Churchill et son équipe de l'Université de Duke ont étudié les membres supérieurs de A.sediba, notamment les humérus, radius, ulna, scapula, clavicule et manubrium les plus intacts découverts à ce jour sur les premiers homininés, tous appartenant au même individu. A l'exception du squelette de la main, les membres supérieurs des homininés de Malapa présentent une morphologie largement primitive. Ainsi, A.sediba partage avec les autres Australopithèques des longs bras et un torse puissant bien adaptés à la progression dans les arbres et probablement à la suspension, et peut-être même mieux qu'on l'avait supposé pour ce genre d'homininé.
Les fragments de la cage thoracique de A.sediba ont été décrits par Schmidt et son équipe et révèlent un buste médiolatéral étroit similaire à celui des grands singes et différent du torse cylindrique des humains. Combiné aux fragments d'une épaule, l'image morphologique qui en émerge est celle d'un thorax conique avec de grandes jointures aux épaules, donnant l'impression que A.sediba "roule des épaules", une configuration peut-être unique chez les Australopithèques et qui n'aurait pas conduit au balancement caractéristique des bras que présentent les humains quand ils marchent à grand enjambées ou pendant la course. Les éléments moins bien conservés de la partie inférieure du tronc suggèrent un degré d'humanisation médiolatéral se rétrécissant dans la partie inférieure, indiquant une anatomie de la poitrine inattendue qui ne ressemble pas à celle observée plus tard chez l'Homo erectus ou l'Homo sapiens.
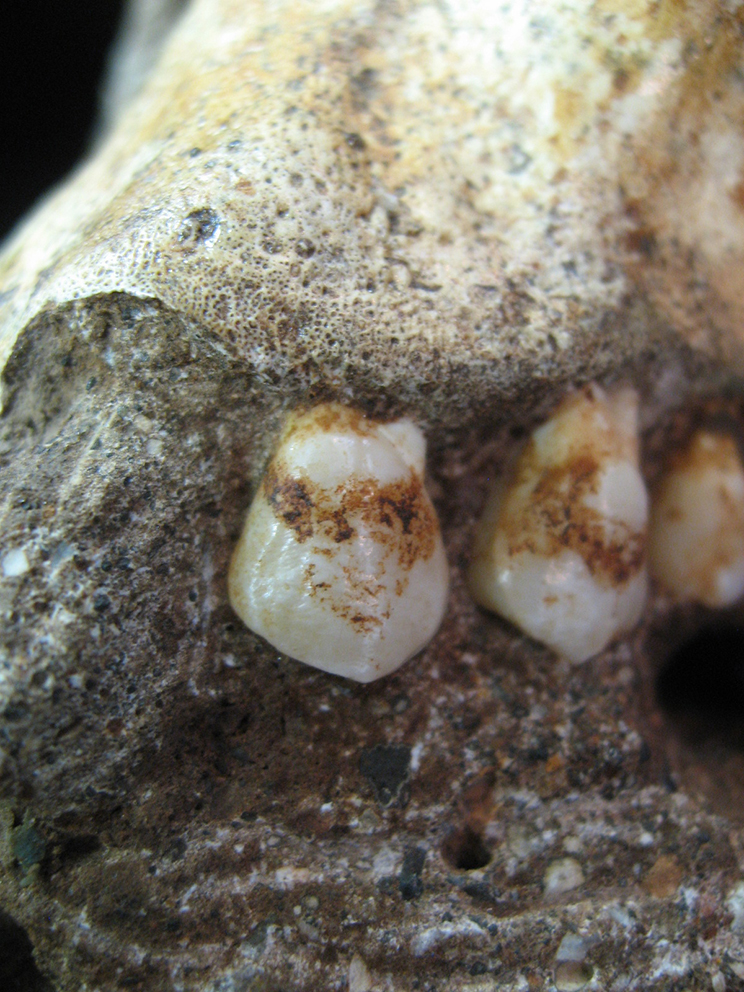
Williams et son équipe ont analysé les éléments des vertèbres cervicales, thoraciques, lombaires et de la région du sacrum et révélé que A.sediba présentait le même nombre de vertèbres lombaires que les humains modernes mais le bas du dos était fonctionnellement plus long et plus flexible. Les indicateurs morphologiques d'une forte courbure lombaire suggèrent que A.sediba hérita à ce sujet de certains traits de l'A.africanus et qu'il ressemblait au squelette de l'homme de Nariokotome (le "garçon de Turkana", un Homo habilis ou un Homo ergaster selon les experts, on y reviendra). DeSilva et son équipe ont étudié les membres inférieures de A.sediba. et proposé une hypothèse biomécanique spécifique sur la façon dont marchait cette espèce. Isolément, les anatomies du talon (petit et qui ressemble à celui du chimpanzé), de la jambe, du genou, de la hanche et du dos sont uniques et curieuses. Mais leur combinaison est cohérente avec la marche bipède avec une gaine hyperpronatée. La combinaison de caractères primitifs et dérivés chez l'Australopithecus sediba indique qu'il s'agit d'une forme transitoire entre la forme adaptée à une vie partiellement arboricole et une forme primaire adaptée à la marche bipède mais dont les jambes et les pieds sont hérités d'une manière de marcher debout encore inconnue. Cela implique que plusieurs formes de bipédisme ont été pratiquées par les premiers ancêtres des homininés. La démarche normale de A.sediba présente une rotation intérieure des genoux et des hanches avec une foulée légèrement tordue et des pieds plats tournés vers l'intérieur pendant que le poids du corps est reporté sur l'extérieur du pied, une posture qui devait sans doute l'aider à marcher au sol. Selon Lee Berger du Wits Evolutionary Studies Institute de l'Université de Witwatersrand d'Afrique du Sud, "c'est un compris parfait pour quelque chose qui a besoin de marcher par terre efficacement sur de longues distances. En même temps, il est tout à fait capable de grimper". L'examen d'un grand nombre d'éléments du squelette d'A.sediba souvent complets et intacts nous laisse entrevoir une espèce d'homininé ayant une anatomie mosaïque présentant des caractéristiques fonctionnelles différentes de celles des autres espèces d'Australopithecus et des premiers Homo. Des mâchoires et des dents riches d'indices L'analyse de la dentition de l'Australopithecus sediba montre que cette espèce est distincte des autres clades d'Australopithèques. En effet, les fragments de mandibules du spécimen MH-2 et d'autres échantillons d'incisives et de prémolaires d'A.sediba jusque là non identifiés ont été étudiés par De Ruiter et son équipe. Comme on l'a déjà constaté avec d'autres parties du squelette et du crâne de cette espèce, ces fragments de mandibules partagent des similitudes avec ceux d'autres Australopithèques mais sont différents de ceux de A.africanus, à la fois par leur taille, leur forme et leur trajectoire de croissance. Ces résultats renforcent l'idée que A.sediba est génétiquement différent des espèces d'Australopithèques d'Afrique de l'Est et distinct de l'A.africanus vivant en Afrique du Sud. En fait, les mandibules de A.sediba partagent certains caractères hérités d'une population comprenant quatre fossiles du genre Homo. Ces résultats ont des implications sur la manière de considérer la phylogénie des homininés tout le long de la fin du Pliocène et suggère la possibilité que A.sediba et peut-être A.africanus, ne descendent pas de la lignée de l'A.afarensis. L'analyse de son paléoenvironnement et de sa dentition viennent supporter cette hypothèse.
L'analyse des résidus prélevés dans le tartre des dents d'A.sediba indique la présence de phytolithes (microfossiles végétaux) contenant des cristaux de silice suggérant qu'il utilisait ces plantes comme moyen d'auto-défense. A.sediba se nourrissait avant tout de fruits, de différentes espèces de feuilles et d'herbes (ce qu'on appelle les aliments C3) et de tiges tendres plutôt que d'aliments plus lourds de type C4 (qu'on retrouve aussi sur le gisement) comme en consommaient les autres homininés de cette époque qui se nourrissaient d'herbes tropicales et de carex (laîches). Les différentes preuves montrent que A.sediba recherchait des aliments de type C3 typiques des galeries forestières entourées de plantes grasses de type C4. L'analyse isotopique du carbone de ses dents montre aussi que A.sediba s'est nourri d'aliments similaires à ceux qu'on trouve aujourd'hui dans la savane et dont se nourrissent les chimpanzés ou les girafes. De plus, peu avant son décès A.sediba a consommé des aliments plus durs comme des noix et même un aliment auquel on ne pensait pas, de l'écorce comme l'explique l'article publié par Lee Berger et son équipe dans "Nature" en 2012. Comme le souligne Berger, de nombreux primates mangent des noix tombées des arbres lorsque les fruits deviennent difficiles à trouver. On peut imaginer que A.sediba dut s'en contenter à une époque où le climat s'est asséché forçant les homininés à se rapprocher des points d'eau. Ces noix pourrait renforcer le scénario selon lequel "nous avons peut-être trouvé une communauté qui vivait juste au moment où son environnement est devenu incroyablement stressant", conclut Berger. Toutefois, une étude publiée en 2016 par Justin A. Ledogar et son équipe à propos des dommages dentaires microscopiques observés sur les deux spécimens indique que A.sediba ne disposait pas de mâchoires suffisamment robustes pour manger régulièrement des aliments durs comme des noix ou des graines (pas plus que la mâchoire des hommes modernes). Cette étude offre un bon exemple de la manière dont les outils modernes de l'ingéniérie peuvent être utilisés pour répondre aux questions évolutionnaires. Dans ce cas, ils permettent d'avoir une meilleure compréhension du squelette facial et ce qu'il peut nous révéler sur le régime et le style de vie des ancêtres des hommes et des primates. Ainis, selon Ledogar, "si certaines populations d'Australopithèques ont évolué pour s'adapter et maximiser leur capacité à mordre, d'autres dont A.sediba, ont évolué dans le sens opposé" et certaines de ces populations ont finalement donné naissance à l'espèce Homo. Selon David Strait qui collabora à cette étude et étudia l'adaptation écologique des premiers humains ainsi que l'origine et l'évolution de la bipédie, "une clé pour comprendre les origines de notre genre est de réaliser que les facteurs écologiques ont dû perturber les comportements alimentaires et les régimes des Australopitèques. Le régime alimentaire a probablement joué un rôle clé dans l'origine de l'espèce Homo". En conclusion, la consommation d'aliments durs n'a vraisemblablement pas conduit la sélection naturelle à favoriser les espèces limitées par leurs capacités à mordre. Cela signifie que le type de nourriture était très important pour la survie de A.sediba qui devait trouver le moyen de se nourrir facilement sans disposer d'une puissante mâchoire. Des fractures suspectes Aucun outil n'est associé aux fossiles de A.sediba. Toutefois quelques ossements présentent des signes de fractures prouvant qu''ils n'étaient pas seulement des bipèdes chasseurs-cueilleurs mais également des proies de prédateurs, lions et autres léopards et peut-être occasionnellement la cible d'autres Australopithèques ou des premiers Homo. En effet, à l'image du chimpanzé, le fait que A.sediba présente des mains dont le pouce est opposable supporte l'hypothèse qu'il était capable de façonner et d'utiliser des outils. Mais rien ne prouve qu'il utilisa des pierres taillées pour chasser ou se battre bien que cette invention rappelons-le remonte à 3.4 millions d'années, à l'époque de l'A.afarensis. Il n'existe pas non plus de preuves attestant que A.sediba aurait pu se nourrir de viande. S'il y a peut-être goûté, il ne s'y est pas adapté et n'est donc pas un omnivore ni un prédateur. Ces faits prouvent que la théorie avancée dans les années 1950 par Raymond Dart à propos des origines africaines de l'homme est correcte, en revanche sa théorie selon laquelle nos ancêtres ont survécu grâce à leur agressivité n'est pas démontrée chez A.sediba. Un ancêtre ou un cousin ? Comment expliquer la présence de A.sediba il y a plus de 1.9 million d'années alors même que l'Homo habilis apparut voici 2.4 millions d'années ? La théorie des équilibres ponctués pourrait aider les évolutionnistes à expliquer la survie de A.sediba à cette époque "récente". Sa survivance pourrait signifier que A.sediba serait apparu peu avant ou en même temps que l'Homo habilis. La chronologie serait alors la suivante : A.africanus (3.5 à 2.5 millions d'années) → A.sediba (2.4 à 1.75 million d'années) → H.habilis (2.4 à 1.3 million d'années). Dans ce cas, A.sediba serait notre cousin et non pas notre ancêtre. Encore faut-il le démontrer. Après dix ans de recherches, les paléoanthropologues ont fini par trouver une preuve temporelle que ce cousin éloigné de "Lucy" n'est probablement pas le progéniteur de l'espèce humaine. Cette annonce fut publiée dans la revue "Science Advances" en 2019 par deux chercheurs de l'Université de Chicago, le postdoctorant en paléontologie évolutionnaire Andrew Du et le paléoanthropologue éthiopien Zeresenay Alemseged, déjà connu pour avoir découvert Selam (DIK-1-1) surnommée "la petite Lucy". La conclusion des chercheurs, même avec leur prudence probabiliste fut accueillie avec scepticisme par certains spécialistes, principalement parce que les chiffres ne semblaient pas concorder. Faisons le calcul. D'une part, en raison de l'âge des fossiles de A.sediba, l'espèce existait déjà environ 200000 ans avant le premier représentant incontesté du genre humain, l'Homo erectus. Toutefois, deux autres membres contestés du groupe, l'Homo rudolfensis et l'Homo habilis sur lesquels nous reviendrons, coexistaient probablement en même temps que A.sediba. D'autre part, un autre élément de preuve qui remet sérieusement en cause l'idée que A.sediba serait l'ancêtre du genre humain (Homo sp.) fut mis au jour en 2015, avec la découverte à Ledi-Geraru dans l'Afar, en Éthiopie, d'un os de la mâchoire et des dents d'un individu appartenant avec certitude au genre Homo, mais qui date de 2.8 millions d’années (cf. B.Villmoare et al., 2015). L'espèce fossile n'a pas encore été nommée, mais selon Du et Alemseged, son existence remet en cause l’affirmation selon laquelle A.sediba serait notre ancêtre. S'il l'était, cela voudrait dire que le premier ancêtre de l'Homme survécut 800000 ans après l'émergence de son propre descendant. Même si en théorie ce n'est pas impossible, les chercheurs ont rapidement admis que c'est extrêmement peu probable et ont tout mis en oeuvre pour le démontrer.
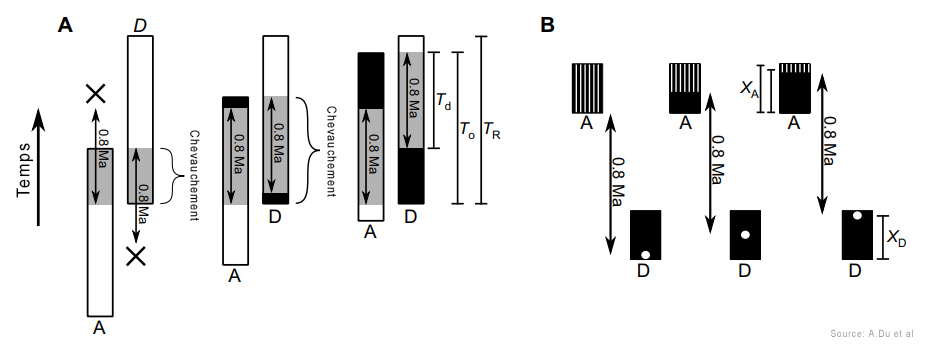
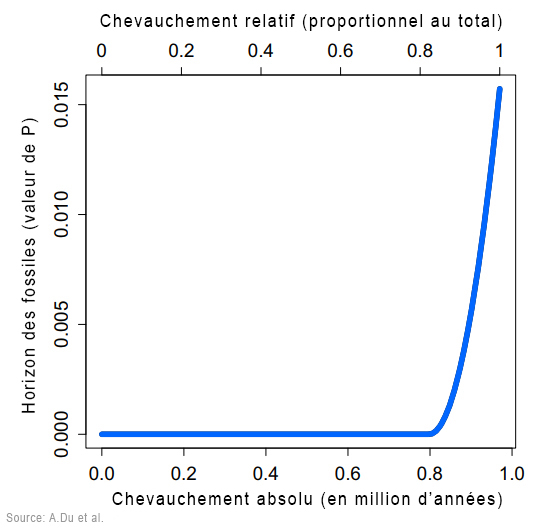
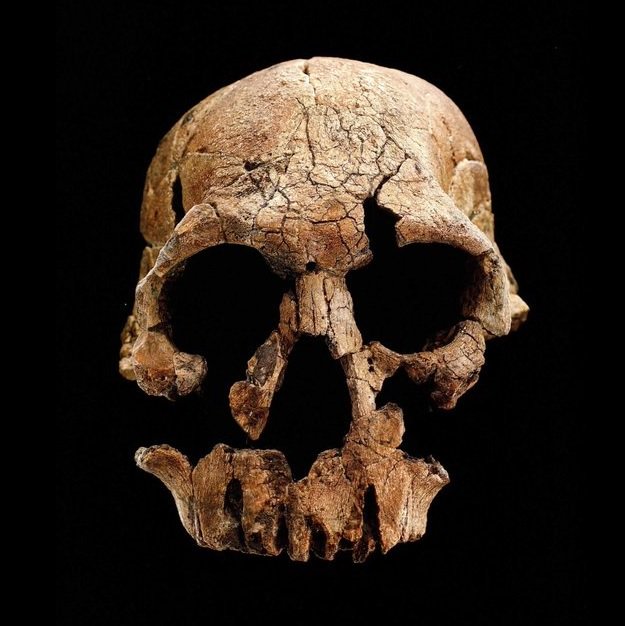
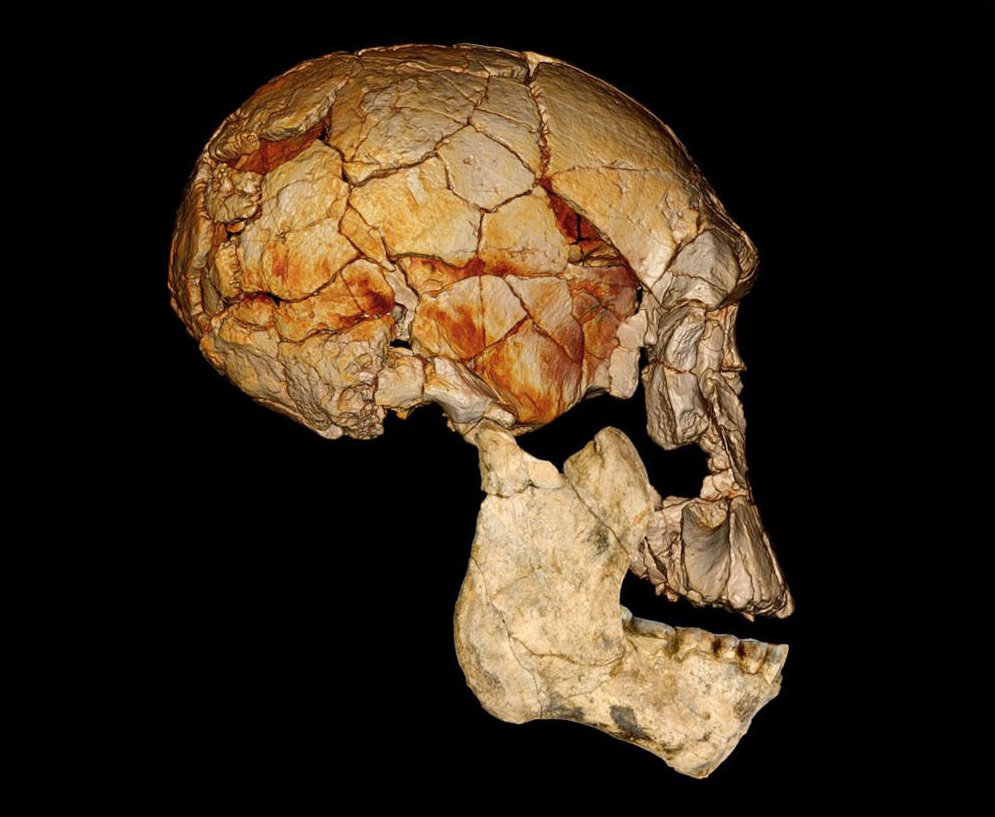
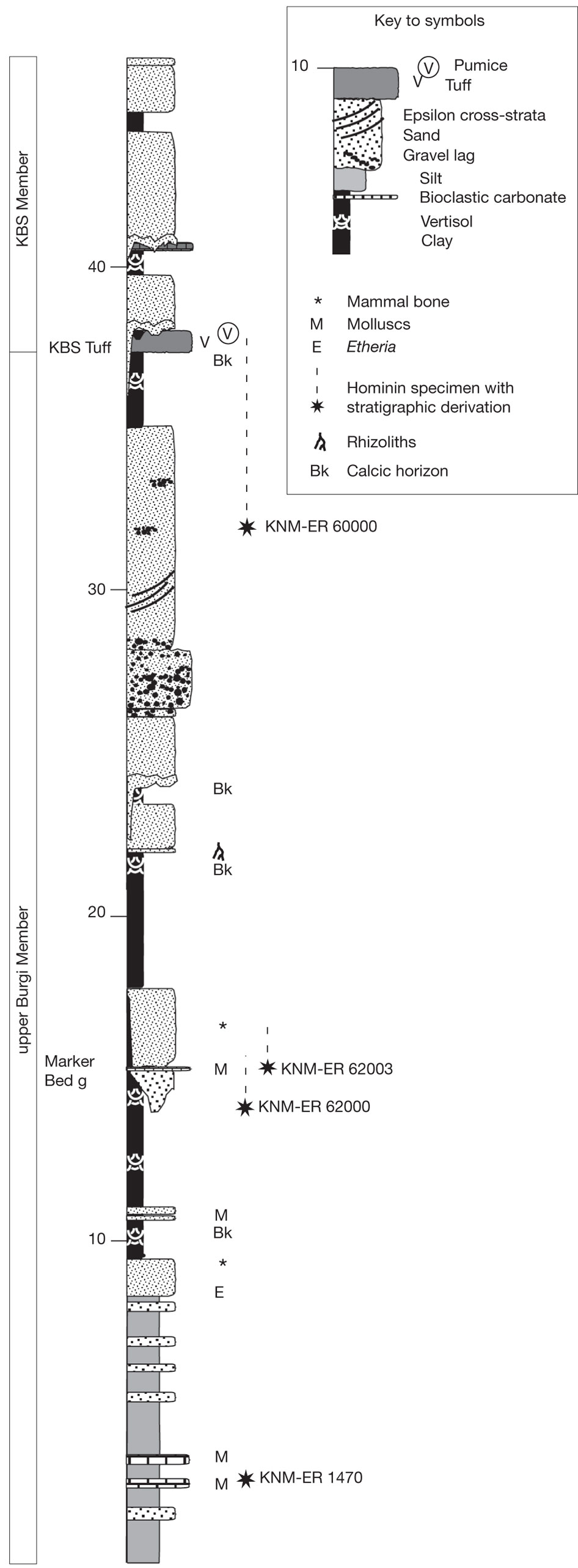
D'abord, Du et Alemseged ont dû formuler une hypothèse testable. Ils ont supposé qu'à un moment de leur histoire, une sous-population de A.sediba fut isolée du reste de l'espèce et, probablement dans un environnement aux contraintes très différentes, évolua en une créature distincte. Dans le jargon, ils supposaient que le genre Homo fut "créé cladogénétiquement" - c'est la multiplication des espèces - à partir de A.sediba. Selon les chercheurs, sans cette hypothèse "cette analyse serait inutile et l'argument voulant que A.sediba soit l'ancêtre du genre Homo serait illogique (car l'horizon des fossiles de A.sediba est postérieur au premier horizon des Homo)". Les chercheurs ont ensuite évalué les probabilités de découvrir de nouveaux fossiles des deux espèces afin de calculer la probabilité que l'extinction éventuelle de A.sediba soit postérieure à celle de son descendant présumé, le premier du genre Homo pour l'instant anonyme. L'analyse a révélé que la probabilité que A.sediba soit un ancêtre direct était de 0.09%, ce qui est très faible. Mais jamais trop prudent, en guise de vérification supplémentaire, Du et Alemseged ont vérifié dans la littérature paléoanthropologique s'il existait d'autres cas où des ancêtres homininés survécurent jusqu'à la période de leur descendance. En examinant 28 cas potentiels d'homininés issus de la descendance ancestrale, ils ont trouvé un exemple : un fossile d'Homo erectus datant d'environ 100000 ans après l'émergence de son descendant, l'Homo antecessor. Selon Du, "encore une fois, nous constatons qu'il est possible que le fossile d'un ancêtre soit postérieur à celui de son descendant. Mais 800000 ans, c'est assez long". En conclusion, les auteurs suggèrent que le poids de la preuve montre que le véritable ancêtre de la lignée Homo est un autre australopithèque, l'Australopithecus afarensis, espèce à laquelle appartient le fameux squelette de Lucy. Si cette question semble résolue, à ce jour on ignore encore l'extension géographique et combien de temps survécut A.sediba. On ignore également s'il cohabita avec des espèces humaines. Peut-être le confirmera-t-on un jour. Par conséquent, les similitudes entre A.sediba et H.habilis (tels que le pelvis et le crâne ressemblant à celui des humains) pourraient s'expliquer soit par l'existence d'un ancêtre commun proche des humains dans la lignée ancestrale des Australopithèques, qui resterait à découvrir, soit dans une convergence voire un métissage. En effet, bien qu'il soit plus difficile de le démontrer pour ces époques reculées, il est possible que le métissage ait joué un rôle dans l'adaptation, la mutation et la prolifération des espèces en leur apportant un avantage compétitif supplémentaire. Après tout, depuis l'époque de Lucy le climat et la végétation étaient en train de changer, les prairies ont envahi le paysage et ces changements ont pu affecter les deux espèces apparentées qui partageaient le même habitat. Les études à venir devraient nous en dire plus. L'Homo rudolfensis : 1.95 - 1.75 million d'années En 1972, au cours d'une expédition paléontologique dirigée par Richard Leakey à Koobi Fora à l'est du lac Rudolf (aujourd'hui rebaptisé lac Turkana), au Kenya, Bernard Ngeneo découvrit sous la couche de tuf les fragments d'un crâne fossile qui sera baptisé "KNM-ER 1470" (ER 1470 en abrégé). L'équipe mit également à jour un fémur complet et des fragments du second fémur, d'un tibia et d'une fibule distale (os de la jambe).
Meave Leakey et Bernard Wood mettront plusieurs semaines pour reconstituer les 150 fragments du crâne de KNM-ER 1470, révélant un crâne large avec une face longue, large et plate. Alors que les racines des dents supérieures indiquaient clairement qu'il avait de larges dents, le crâne ne présentait pas les détails des stuctures musculaires massives caractéristiques des Australopithecus robustus. Sans disposer d'autres échantillons, Richard Leakey pensa que cet holotype était vieux de 2.9 millions d'années et le classifia comme "une espèce Homo indéterminée", Homo sp. Une naissance difficile Après une nouvelle datation, les scientifiques déclarèrent que le crâne était âgé de 1.9 million d'années, et donc contemporain de l'Homo habilis. La communauté scientifique pensa bien faire en classant KNM-ER 1470 parmi les Homo habilis bien que sa mâchoire et ses dents ne rentraient pas dans les limites de variabilités acceptables pour ce taxon. Cette découverte fut à l'origine d'un débat qui dura plus de quarante ans durant lesquels les paléoanthropologues discutèrent de la taxonomie et l'origine des fossiles découverts depuis 1959 dans l'est de l'Afrique et attribués à l'Homo habilis.
Selon le point de vue des experts, les fossiles découverts à Koobi Fora au Kenya (préfixe ER), à Olduvai en Tanzanie (préfixe OH) et à Hadar dans la vallée de l'Omo en Ethiopie (préfixe Omo) appartiennent pour les uns à l'Homo habilis, pour les autres à deux taxons différents dont une nouvelle espèce comme le pensait Leakey, certains n'y voyant que des sous-espèces d'Australopithèques. A charge des défenseurs de la thèse de l'Homo habilis, il paraît difficile d'imaginer que tous ces fossiles dispersés sur une période d'environ un million d'années (entre -2.45 et -1.55 million d'années) appartiennent à un seul et même taxon; la sélection naturelle a nécessairement fait évoluer ces individus qui présentent de fait des différentes morphologiques significatives les éloignant parfois trop pour appartenir à la même espèce. Que nous révèlent les fossiles de KNM-ER 1470 ? Selon les chercheurs du CNRS, sa voûte crânienne est plus épaisse que celle de l'Homo habilis. Son volume endocrânien est estimé entre 650 et 750 cm3 soit 1.5 fois supérieur à celui des Autralopithèques découverts en Afrique du Sud et légèrement supérieur à celui de l'Homo habilis (600-700 cm3). A consulter : Le corps humain, René St-Jacques Structure anatomique du crâne, Imaios
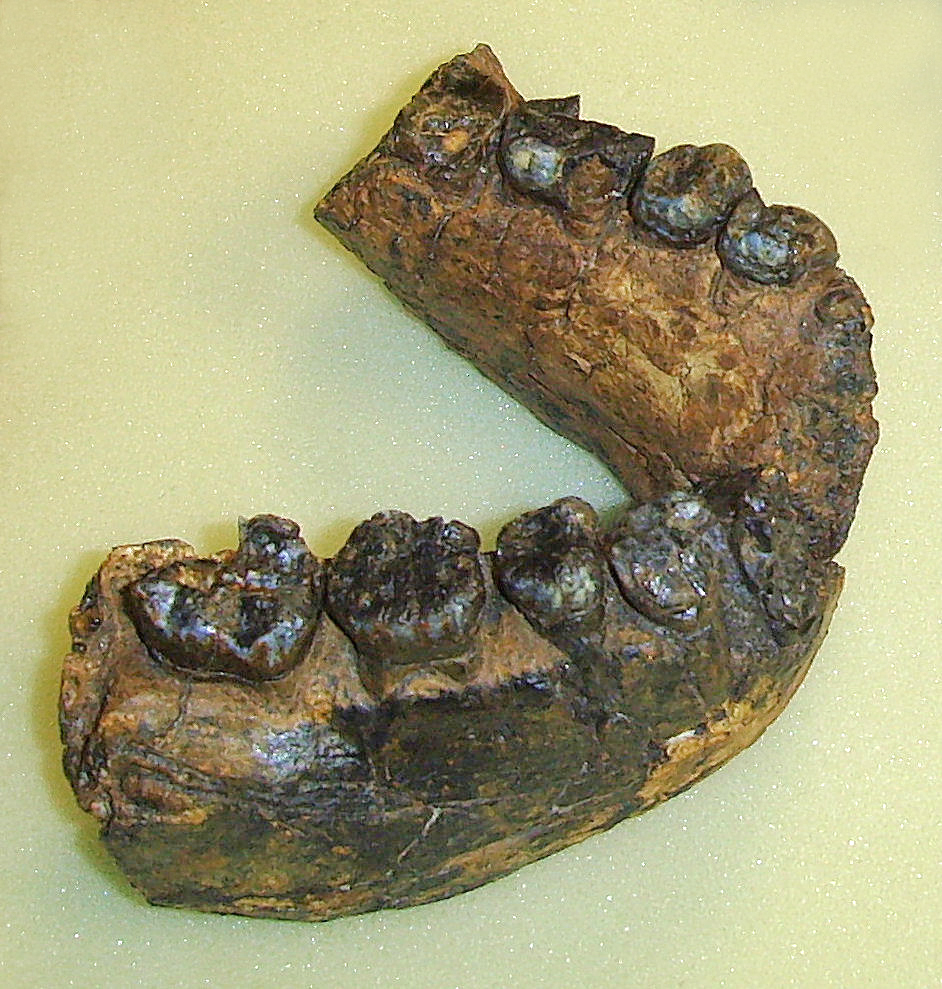
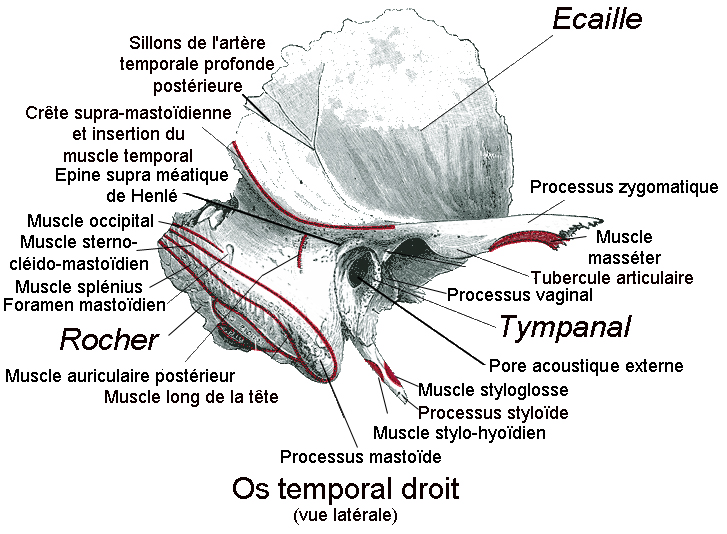
Sa mâchoire est épaisse et robuste, le rapprochant de la lignée des Paranthropus (à laquelle appartient P.robustus alias l'Australopithecus robustus) et indique qu'il consommait des aliments carnés. Ses orbites sont extrêmement hautes et ont une forme carrée alors qu'elle est rectangulaire avec un allongement supéro-inférieur chez l'Homo habilis. Il n'y a pas de dépression à l'arrière de la région orbitaire ni de dépression postorbitaire latérale, contrairement à l'Homo habilis. La forme de l'écaille de l'os temporal est haute et arrondie, alors qu'elle est basse et triangulaire chez l'Homo habilis. Les crêtes mastoïdiennes et supramastoïdiennes situées sur l'os temporal (à hauteur de l'arcade zygomatique), sont jointives alors qu'il n'y a pas de jonction chez l'Homo habilis. Le torus supra-orbitaire (l'arcade sourcilière) est développée plus médialement que latéralement, l'ouverture nasale est très étroite et le torus occipital est absent alors qu'il présente un développement médial chez l'Homo habilis. Le squelette étant très incomplet, les paléoanthropologues ont étudié les autres spécimens datant de la même époque et découvrirent que certains présentaient ces mêmes différences et bien d'autres avec l'Homo habilis, comme par exemple un menton plus rectangulaire que celui de l'Homo habilis qui présente une convexité médio-latérale.
Par bien des côtés, tous ces spécimens sont plus évolués que les Australopithèques mais sont également différents dans leurs détails des Homo habilis. Finalement, en 1978, après un nouvel examen le paléoanthropologue russe Valeri Alexeev considéra que KNM-ER 1470 appartenait à une nouvelle espèce d'homininé baptisée Pithecanthropus rudolfensis par référence au lac près duquel elle fut découverte. Toutefois, en 1986 Alexeev rebaptisa le genre en Homo rudolfensis qui fut accepté par la communauté scientifique. Entre 2007 et 2009, soit près de 40 ans après la découverte de KNM-ER 1470, Meave Leakey et son équipe découvrirent dans la même région sédimentaire entourant le lac Turkana mais dans un strate un peu moins profonde, trois nouveaux fossiles âgés entre 1.78 et 1.95 million d'années. Il s'agissait d'une mandibule complète (KMN-ER 60000), des fragments d'une face et d'une mâchoire juvénile (KMN-ER 62000) et d'un fragment de mandibule contenant quatre dents (KNM-ER 62003). Leur morphologie était compatible avec l'arc dentaire de KNM-ER 1470. Cette découverte confirmait les caractéristiques uniques du visage de KNM-ER 1470 et qu'il s'agissait bien d'une espèce distincte de l'Homo erectus et de l'Homo habilis. Ces découvertes appuyent également la thèse de l'existence simultanée de plusieurs espèces d'homininés en Afrique de l'Est il y a 2 millions d'années. En effet, les fossiles de l'Homo rudolfensis confortent l'idée d'une cohabitation avec l'Homo habilis. Il vivait également à la même époque que l'Homo ergaster (voir page 8), représenté par le crâne KNM-ER 3733 âgé de 1.7 million d'années et par le "garçon de Turkana" (KNM-WT 15000) âgé d'environ 1.6 million d'années et un autre spécimen découvert en Chine. Selon Bernard Wood, il est même "parfaitement possible" qu'il y ait eu des interactions entre ces différentes espèces. En 2007, une équipe d'anthropologues dirigée par Timothy Bromage de l'Université de New York, déjà connu pour ses reconstructions paléoanthropologiques numériques (basées sur la technologie de la tomographie comme le scanner ou CT scan), reconstruisit le crâne de KNM-ER 1470 en lui ajoutant la mâchoire inférieure du spécimen KNM-ER 60000. La reconstruction ressemblait plus au crâne d'un singe qu'à celui d'un homme et sa capacité crânienne fut réduite de 752 cm3 à environ 526 cm3, ce qui n'était pas très flatteur pour ce premier représentant humain. Mais d'autres squelettes lui donneront finalement des proportions plus humaines. Une espèce isolée et sans descendance Tous les spécimens d'Homo rudolfensis sont omnivores, de petite taille (~1.25 m) et leur poids adulte se situe entre 50 et 52 kg. L'Homo rudolfensis vivait en Afrique de l'Est (Ethiopie, Kenya, Tanzanie) ainsi qu'au Malawi (site de Uraha) et en Afrique du Sud (sites de Drimolen, de Swartkrans et de Sterkfontein). Une mandibule fut également découverte en Chine (site de Longuppo) et serait apparentée avec l'Homo habilis ou l'Homo erectus. L'Homo rudolfensis présente des caractéristiques spécifiques mais déjà marquées par une variabilité qu'on retrouvera pas la suite chez l'Homo erectus et l'Homo sapiens, notamment la courbure saggitale de l'os pariétal plus ou moins prononcée (faible chez les homininés), l'angle de courbure externe de l'os occipital relativement ouvert et variable (proche de l'Homo sapiens), des incisives assez larges (plus larges que celle de l'Homo erectus) et quelques éléments de son squelette (os des pieds, des mains, la clavicule) très similaires à celles de l'Homo sapiens. Néanmoins, à ce jour il est très difficile de déterminer avec précision qu'elle est son ancêtre direct. On peut imaginer qu'il hérita certaines caractéristiques des Australopithèques mais duquel d'entre eux, A.anamensis, A.africanus ou même d'un autre taxon, nul ne le sait. Une chose est sûre, l'Homo rudolfensis ne survécut que 200000 ans puis son espèce s'est éteinte. Ceci dit, le travail d'enquête des paléoanthropologues n'est pas achevé et chaque année nous apporte son lot de découvertes. Prochain chapitre
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||










{kind=link}