|
|
|
L'origine et l'avenir de l'Homme
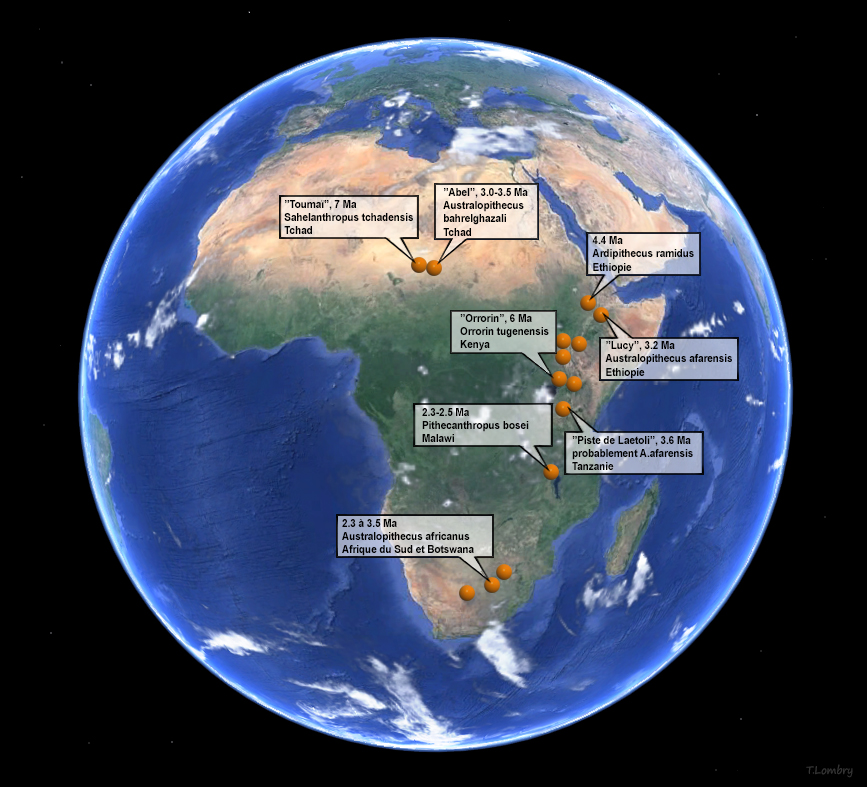
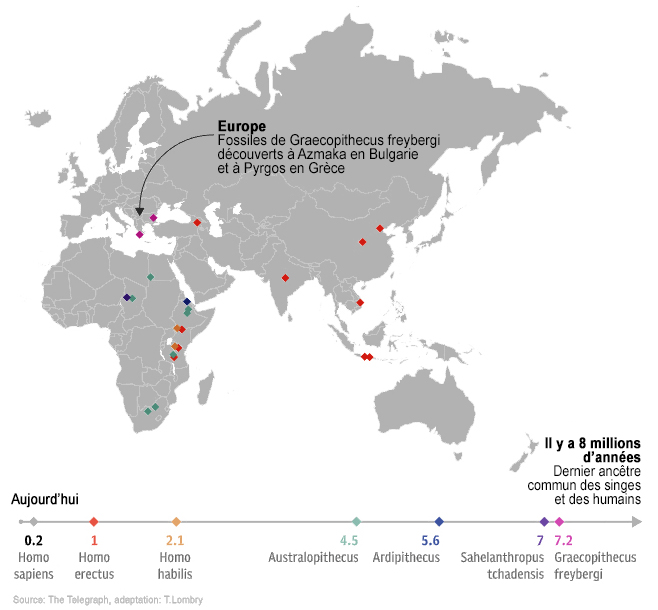
Le berceau de l'humanité (III) De toute évidence, si on se réfère à la localisation des plus anciens fossiles d'homininés (la ligné des hominidés conduisant au genre Homo comprenant notamment les Autralopithèques) découverts à ce jour, l'origine de l'homme est unique et se situe quasi certainement en Afrique tropicale[1]. En effet, bien que des fossiles d'homininés existent en Asie, ceux-ci ne remontent qu'à 2 millions d'années alors qu'on retrouve en Afrique des fossiles préhumains isolés remontant à 7 millions d'années. Jusqu'en 2001, il était généralement admis que le berceau de l'humanité se situait dans l'Est africain, à l'est du Rift Albertin, le long de la "corne" qui va du Kenya à l'Ethiopie. Mais comme on le voit sur la carte présentée à droite, la découverte de Toumaï et d'Abel plus au nord, au Tchad, rend cette théorie caduque; l'East Side Story proposée en 1983 par le paléoanthropologue Yves Coppens doit être sinon abandonnée du moins revue et corrigée. On y reviendra. Quant à la "théorie de la Savane" qui prétend que les préhumains se sont redressés suite à l'assèchement du climat, l'ouverture des forêts et l'extension de la savane qui s'ensuivit, la découverte du squelette d'Ardipithecus ramidus démontre que cet hominidé tout en étant arboricole se tenait déjà sur ses deux pieds. Vu que de nombreux animaux se déplacent en utilisant uniquement leurs pattes arrières, cette convergence évolutive suggère que ce n'est donc pas uniquement le changement climatique qui serait à l'origine de la bipédie de l'homme mais probablement plusieurs facteurs. On y reviendra également. Que nous soyons tous d'origine africaine est certainement correcte mais la distribution de nos ancêtres sur le sol africain apparaît aujourd'hui plus étendue qu'on l'imaginait, couvrant plus de 5000 km, et impose de nouvelles investigations. L'enquête sur nos origines continue. Rappelons tout d'abord quelle est la structure globale de notre arbre phylogénique. L'arbre phylogénique de l'humanité Nous avons vu dans le tableau présenté en introduction que l'homme moderne appartient à la superfamille des hominoïdes, en résumé c'est un grand singe qui eut la chance de développer ses facultés intellectuelles et la bipédie et devint de ce fait un genre à part entier à l'origine de nombreuses espèces d'homininés qui finit par acquérir la faculté de parler et d'écrire. Citons brièvement les principaux représentants de cet arbre phylogénique à partir de la super-famille des Hominoïdes.
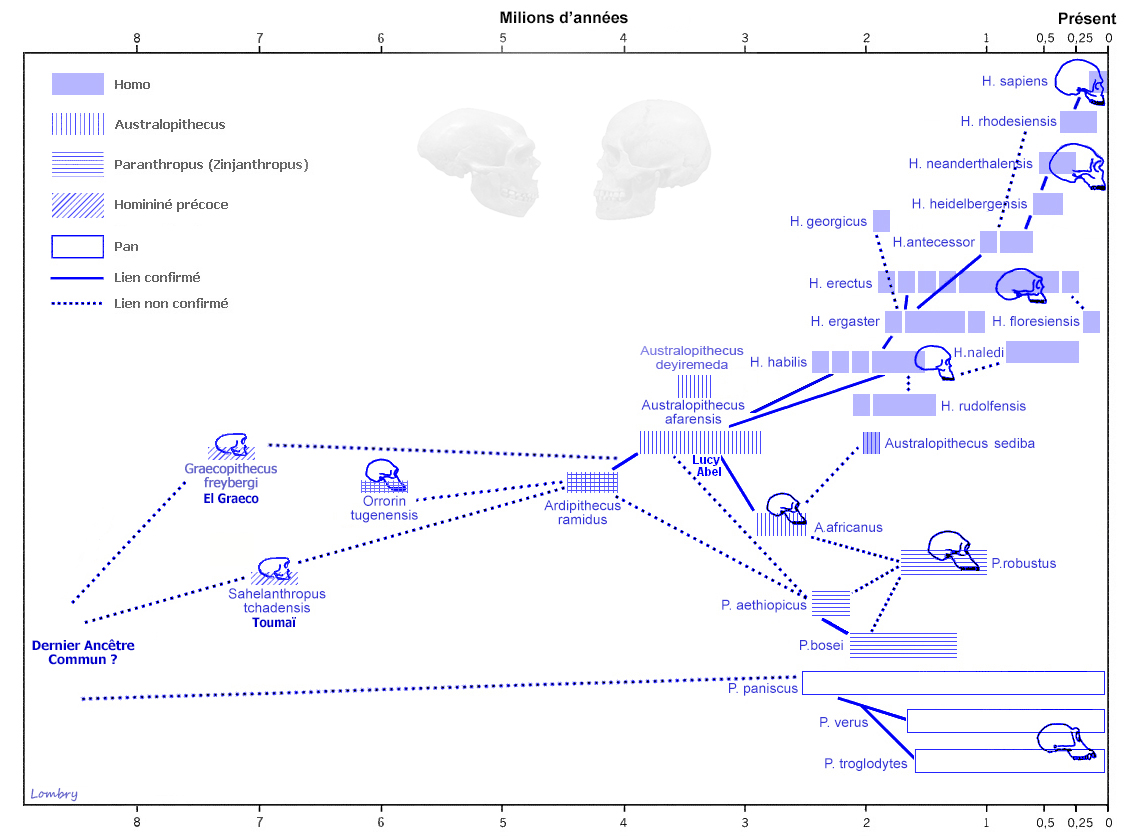
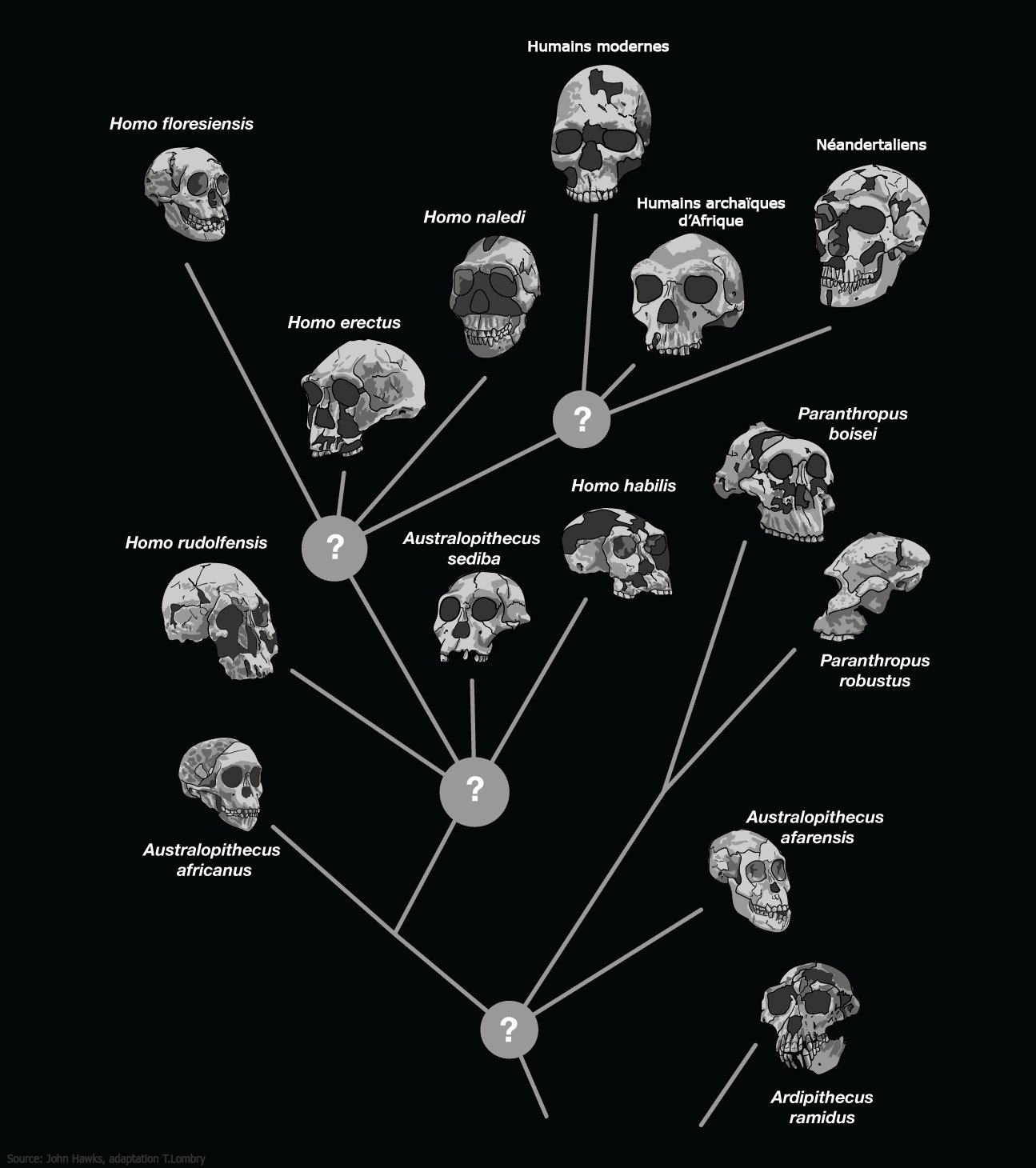
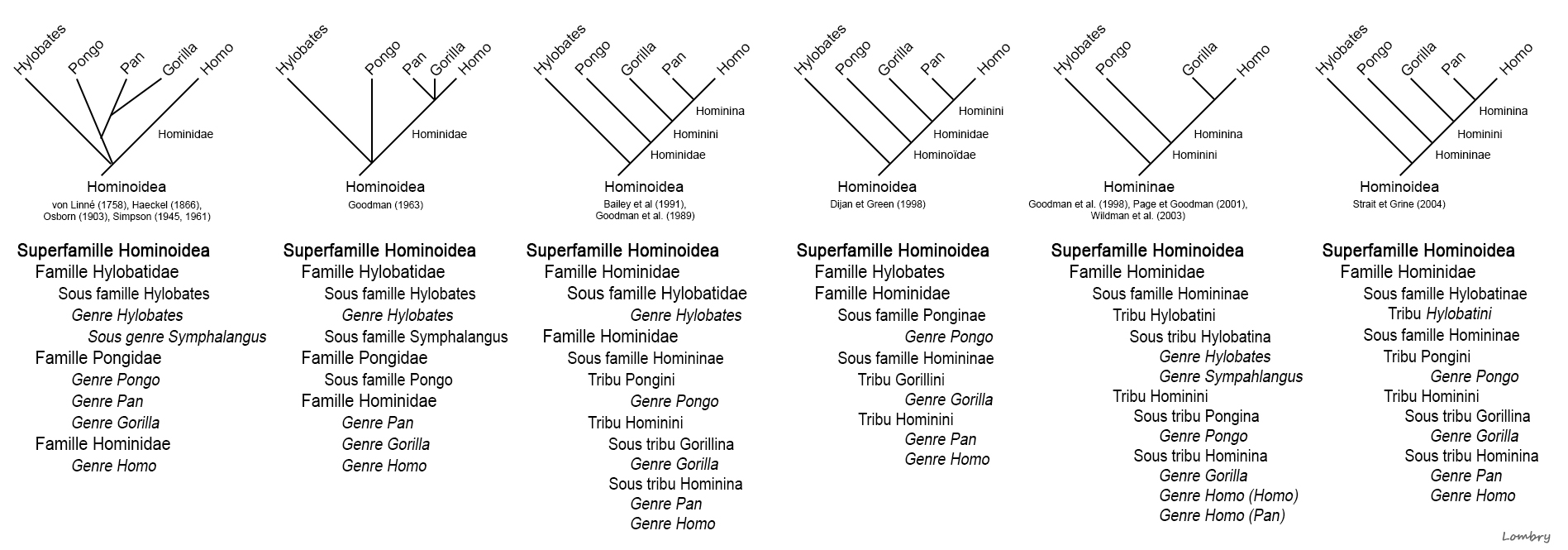
Les fossiles datant de plus de 15 ou 20 millions d'années révèlent la présence d'une dizaine de genres d'hominoïdes que les scientifiques ont divisé en deux grandes familles : les Hylobatidés et les Pongidés/Hominidés. Ces familles sont représentées par 9 genres qui ont donné naissance à plus de 20 espèces. La première famille regroupe les singes Hylobatidés. Ils sont les ancêtres des Hylobates dont descend le gibbon et des Symphalangus dont descend le siamang. La seconde famille regroupe les singes Pongidés et les Hominidés. Les Pongidés sont les ancêtres du genre Pongo (l'orang-outan dont il existe deux espèces) tandis que les Hominidés ont évolué en 2 sous-familles : celle des Gorillinés à l'origine de deux espèces de gorilles (G.gorilla et G.beringei) et les Homininés. Les Homininés ont donné naissance à deux grands genres : Pan (à l'origine de P.troglodytes, l'ancêtre du chimpanzé, et de P.paniscus, l'ancêtre du bonobo) et une branche qui se divisa ensuite en 4 genres : Paranthropus (P.robustus, P.aethipicus et P.bosei), Kenyathropus (K.platyops), Australopithecus (A.anamensis, A.deyiremeda, A.afarensis, A.bahrelghazali, A.africanus, etc) et Homo (H.habilis et au moins 15 autres espèces sur lesquelles nous nous attarderons). Toutefois, malgré cette apparente rigueur, les membres des tribus et sous tribus des cladogrammes de l'espèce humaine font toujours l'objet de débats. Si depuis les années 2000 on observe une convergence entre les chercheurs, il existe toujours des divergences d'opinions sur les questions de détails. A voir : Evolution des cladogrammes de l'espèce humaine (extrait du glossaire)
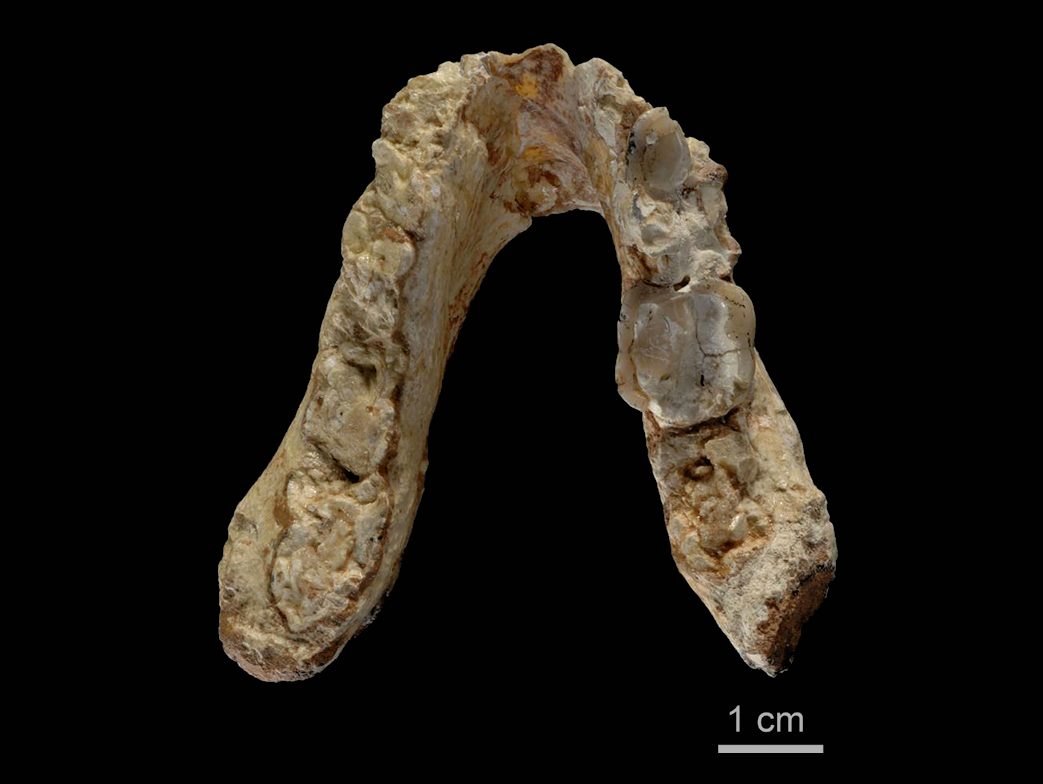
Nous verrons qu'il existe également des espèces d'homininés hybrides entre les singes et les humains tels que le Sahelanthropus tchadensis (Toumaï), Orrorin tugenensis, Ardipithecus ramidus ou encore l'Australopithecus sediba dont la filiation n'est pas encore clairement déterminée du fait que les spécimens sont très peu documentés dans les archives fossiles. Toutes ces espèces vécurent exclusivement en Afrique. A l'exception de quelques rares spécimens comme le Graecopithécus qu'on retrouve dans le sud-est de l'Europe il y a plus de 7 millions d'années (voir ci-dessous), ce n'est que vers 2 millions d'années que leurs descendants ou d'autres espèces plus évoluées ont quitté l'Afrique et peuplé l'Eurasie avant de conquérir le monde. On y reviendra. Dans tous les cas, les traces préhumaines les plus anciennes remontent actuellement à 7.2 millions d'années. Graecopithecus : 7.2 millions d'années Début 2017, une équipe internationale de chercheurs dirigée par la paléoanthropologue et géologue Madelaine Böhme du Centre Senckenberg de l'Université de Tübingen découvrit quelques fragments de mâchoire et une dent en Grèce et en Bulgarie datées de 7.2 millions d'années. Ils appartiendraient à une nouvelle espèce d'hominidé nommée Graecopithecus freybergi. L'annonce de cette découverte fut publiée dans la revue "PLoS One" (cf. art.1 et art.2).
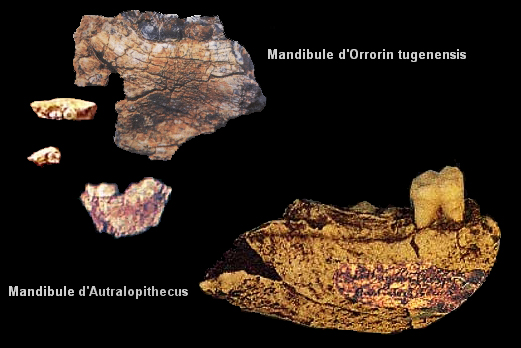
Colin Barras précise dans la revue "New Scientist" que l'holotype de Graecopithecus (ou Grécopithèque) fut découvert en 1944. Il s'agit d'une mandibule fossilisée excavée près d'Athènes dans des circonstances qui restent impprécises en raison de la guerre. A l'époque on pensait qu'elle appartenait à un singe. Suite aux dernières analyses, les chercheurs estiment que la dent découverte en Bulgarie appartient aussi à une espèce de singe qui vécut dans l'est de l'Europe longtemps après la disparition des autres singes du continent. Mais Graecopithecus n'est pas un singe ordinaire, c'est un homininé. Dans la famille taxonomique des Hominidae qui selon les dernières classifications (cf. Goodman et al., 1998 et Wildman et al., 2003) comprend notamment la tribu des Homininés (sous-tribus Pongo, Gorilla, Pan et Homo), les chimpanzés actuels (Pan) sont génétiquement les plus proches des êtres humains (Homo). Se pose alors la question de savoir où vécut le dernier ancêtre commun entre les deux espèces. Jusqu'à la découverte de Graecopithecus, les chercheurs avaient de bonnes raisons de croire en se fondant sur la localisation des fossiles que les deux lignées avaient divergé il y a 5 à 7 millions d'années et que les premiers préhumains sont apparus en Afrique. Or la découverte de fossiles d'homininés précoces en Europe orientale impose que l'on revoit le scénario des premières étapes de l'histoire humaine.
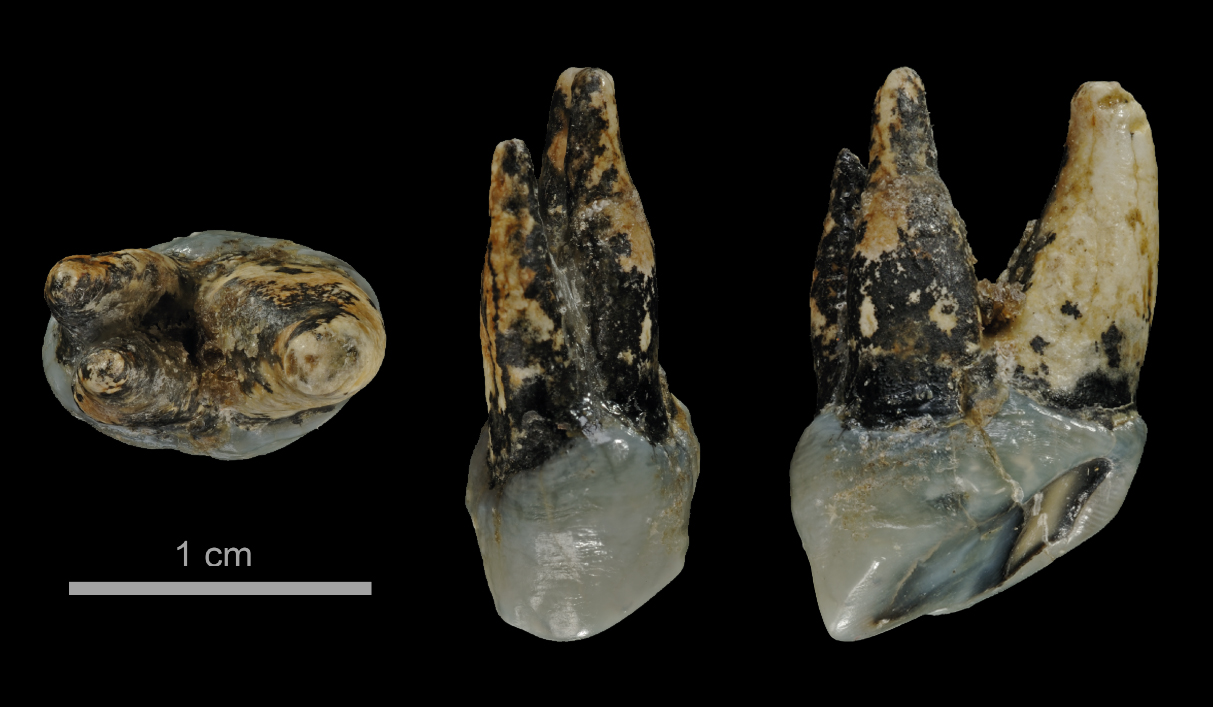
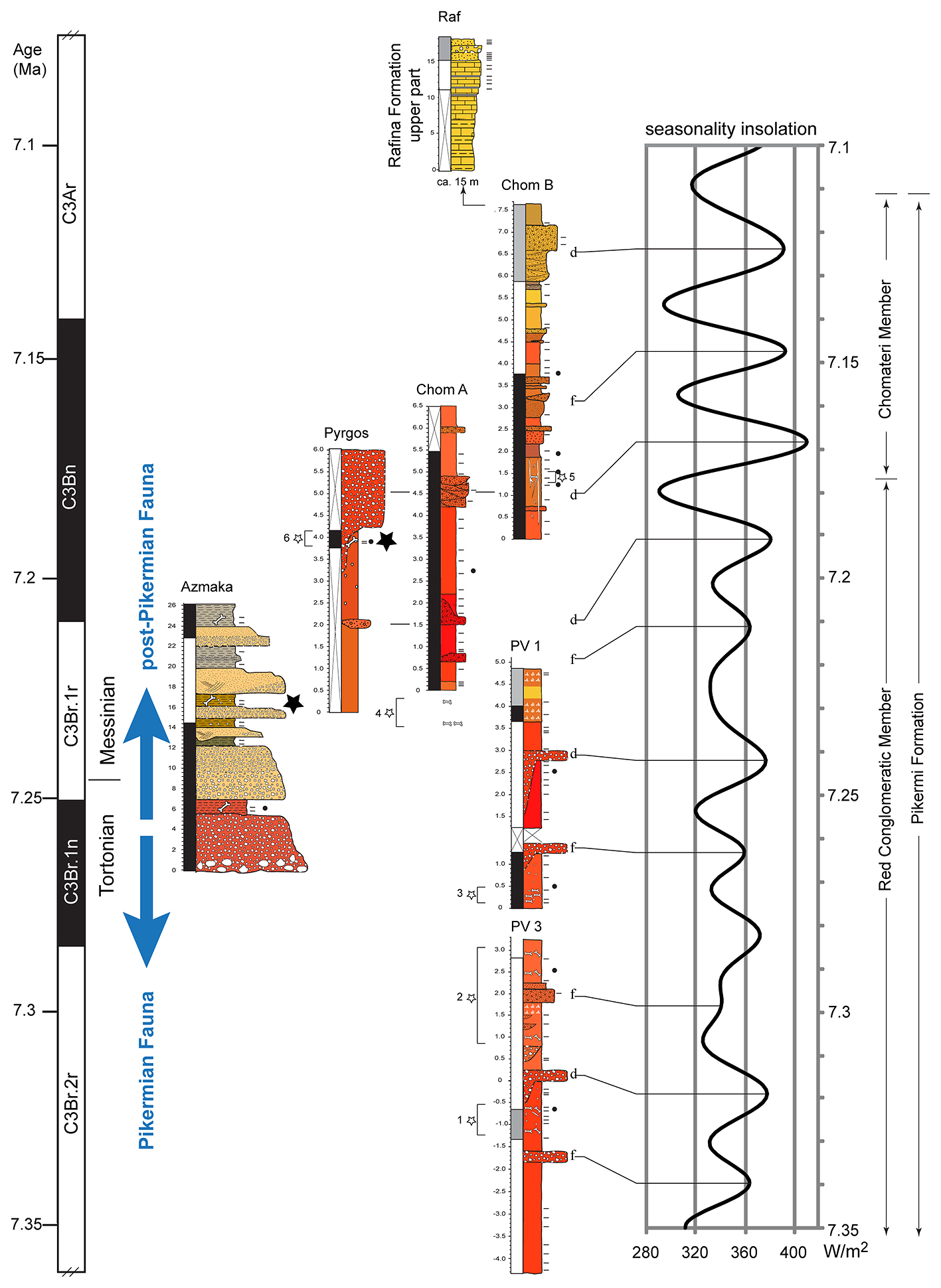
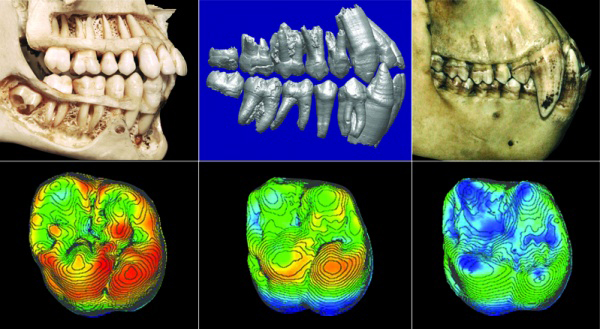
Selon Böhme, "ces différents fossiles indiquent que la scission de la lignée des grands singes et des êtres humains se serait produite plusieurs centaines de milliers d'années plus tôt que prévu et en Méditerranée orientale plutôt qu'en Afrique." Böhme et ses collègues ont analysé les fragments des deux spécimens de Graecopithecus freybergi. En utilisant la tomographie par ordinateur, ils ont visualisé les structures internes des fossiles et ont démontré que les racines des prémolaires étaient presque fusionnées comme on le voit ci-dessus. Selon Böhme,"normalement, chez les grands singes deux ou trois racines sont distinctes et divergentes, alors que les racines de Graecopithecus convergent et sont partiellement fusionnées, une caractéristique typique des humains modernes, des premiers humains et de plusieurs espèces préhumaines dont Ardipithecus et Australopithecus." Les dents de la mandibule du spécimen surnommé "El Graeco" par les scientifiques présentent des caractéristiques racinaires suggérant que l'espèce Graecopithecus freybergi appartiendrait à la lignée préhumaine. En outre, Graecopithecus est plusieurs centaines de milliers d'années plus ancien que Toumaï découvert au Tchad (voir plus bas). Comme on le voit ci-dessous à droite, l'étude stratigraphique des couches sédimentaires des sites grec et bulgare a permis de dater les gisements fossiles de 7.24 et 7.175 millions d'années. Cela nous situe au début du Messinien (cf. la charte stratigraphique et en format PDF), une époque qui se termine par l'assèchement complet de la mer Méditerranée (voir plus bas). Cette datation croisée permet de placer avec certitude Graecopithecus dans la zone méditerranéenne. Les changements environnementaux 1. Une force motrice de la divergence Comme dans la théorie d'Yves Coppens concernant l'émergence de l'homme en Afrique de l'Est sur laquelle nous reviendrons, l'évolution des préhumains a peut-être été motivée par des changements environnementaux spectaculaires. En effet, sur base d'analyses géologiques des grains de poussière, l'équipe de Böhme démontra que le désert du Sahara nord-africain est apparu il y a plus de 7 millions d'années. Quatre longues périodes sèches sont survenues entre 8.75 et 6.25 millions d'années qui ont probablement poussé les animaux à quitter le sud du Sahara et à évoluer sous des climats plus humides, produisant la diversité de créatures qu'on rencontre aujourd'hui dans la savane africaine (cf. M.Böhme et al., 2021). Cet assèchement des sols participa vraisemblablement à l'évolution des hominidés. On reviendra sur le Sahara et les lacs Méga-Tchad et Congo à propos des variations climatiques depuis la Préhistoire. Bien que géographiquement éloignés du Sahara, les limons de couleur rouge découverts dans les strates de Graecopithecus sont très fins et sont très proches des grains de poussière du désert. Une analyse de l'uranium, du thorium et des isotopes de plomb présents dans ces particules leur donne un âge compris entre 0.6 et 3 milliards d'années et les chercheurs en ont déduit qu'ils proviennent d'Afrique du Nord.
De plus, les sédiments poussiéreux sont riches en différents sels. Selon Böhme et ses collègues précités, ces données prouvent pour la première fois que le Sahara s'est étendu il y a 7.2 millions d'années, les orages du désert facilitant le transport des poussières rouges et salées vers la côte nord de la Méditerranée. Ce processus est toujours observable aujourd'hui. Quand le vent du Sahara souffle vers le nord, on retrouve du sable jaune jusqu'en Belgique (cf. les vents régionaux). Cependant, les simulations montrent qu'avec 250 g de sable par mètre carré emporté chaque année, la quantité de poussière à l'époque de Graecopithecus était 10 fois supérieure à celle tombant de nos jours sur le sud de l'Europe et est comparable à la situation actuelle dans la région du Sahel. 2. Savane et stress hydrique en Europe Les chercheurs ont également montré qu'à l'époque de l'extension du Sahara, un biome de savane est apparu en Europe. En utilisant de nouvelles méthodes d'analyses, les chercheurs ont étudié des fragments microscopiques de particules de charbon de bois et de silicates d'origine végétale appelés phytolithes. Beaucoup de phytolithes identifiés proviennent des graminées et en particulier de ceux utilisant la voie métabolique dite de la photosynthèse en C4 (où les premières molécules synthétisées sont des acides organiques à 4 atomes de carbone), celle qui est la plus courante dans les prairies tropicales et les savanes d'aujourd'hui. Les chercheurs ont pu démontrer que la propagation mondiale des graminées en C4 commença il y a 8 millions d'années sur le sous-continent indien, leur présence en Europe étant auparavant inconnue. Selon Böhme, "l'enregistrement phytolithique témoigne de sécheresses sévères et l'analyse du charbon indique des feux de végétation récurrents." Avec l'équipe bulgare, elle a pu reconstruire le biome local d'une savane dans laquelle vivaient des girafes, des gazelles, des antilopes et des rhinocéros aux côtés de Graecopithecus. Notons qu'il s'agit exactement du même décor animalié qu'on retrouvera plus tard sur la "piste de Laetoli" (3.7 Ma) en Tanzanie. Pour Böhme, "la formation naissante d'un désert en Afrique du Nord il y a plus de 7 millions d'années et la propagation des savanes dans le sud de l'Europe a peut-être joué un rôle central dans la répartition des lignées humaines et des chimpanzés." Elle appelle cette hypothèse la "North Side Story", rappelant la thèse d'Yves Coppens connue sous le nom de "East Side Story" sur laquelle nous reviendrons. En résumé, la présence de Graecopithecus en Europe orientale à une époque si reculée remet en question pas mal d'idées établies sur les origines des préhumains. C'est une affaire à suivre dont on reparlera certainement. Et de fait, à peine quelques mois après la publication de cette découverte, des paléoanthropologues annoncèrent la découverte de traces préhumaines à Trachilos en Crète datant de 5.7 millions d'années (cf. page 5). Ceci étant, bien que ce soit spéculatif, il existe peut-être un indice qui va dans le sens de la "North Side Story", c'est l'assèchement de la Méditerranée à la fin du Miocène. Personne ne l'a encore évoqué en relation avec la migration des populations africaines, mais l'idée mérite d'être étudiée. Avis aux chercheurs. Voyons cette théorie. 3. La crise de salinité Messinienne ou comment la mer Méditerranée s'est asséchée À la fin du Miocène, il y a 6 millions d'années, le continent africain et l'Europe sont entrés en collision. Le soulèvement qui en résulta provoqua la fermeture du détroit de Gibraltar. Coupée de sa source d'eau, la mer Méditerranée s'est progressivement évaporée en commençant par la région entourant la Sicile (dont l'Etna culmine à 3350 m au-dessus du niveau actuel de la mer). Le bassin occidental de la Méditerranée fut alors séparé du bassin oriental, les tranformant en d'immenses lacs hypersalés. Suite à la forte évaporation, il ne resta bientôt qu'une étendue boueuse traversée par quelques rivières saumâtres comme nos plages de sable à marée basse. Puis la sécheresse s'accentua et la Méditerranée se transforma en désert. Toute la vie terrestre et aquatique disparut et seuls les oiseaux osaient encore traverser ces étendues stériles par la voie des airs au cours de leurs migrations. Finalement, le fond de la Méditerranée s'est couvert de dépôts de sels atteignant 1 km d'épaisseur. Les chercheurs estiment que durant l'époque Messinienne, le niveau des océans diminua entre 1300 et 2400 mètres (cf. W.Ryan, 2008 et R.Urgeles et al., 2011). A
voir : The evaporating Mediterranean Sea, BBC The Zanclean Megaflood Hypothesis, SciNews, 2018
On estime qu'en moins de 2000 ans, toute la Méditerranée s'assécha et devint une terre inhospitalière, abandonnant derrière elle 1 million de km3 de sels de sodium et de potassium répartis sur plus de 2 millions de km2, ce qui représente des milliards de tonnes de sel. On appelle cette période la crise de salinité Messinienne. Ce sel messinien s'est notamment déposé à une profondeur de 500 mètres sous l'actuelle Sicile, donnant naissance au plus vaste gisement de sel marin du monde, la fameuse mine de sel de Realmonte. On exploite le gisement de sel depuis plus de 50 ans et on pourra encore l'exploiter durant des milliers d'années. Pendant cette sécheresse sans précédent, on suppose que périodiquement de l'eau salée continua de couler car on trouve une alternance de couches de marnes et de gypse sur une épaisseur qui dépasse parfois 300 mètres, comme par exemple sur les côtes de Realmonte qui affichent également de magnifiques falaises de gypse blanc comme on le voit ci-dessous à driote. Par endroit, le flux d'eau fut si important que le gypse a totalement été érodé.
Dans une deuxième phase, il y a environ 5.4 millions d'années et pendant 100000 à 200000 ans, le régime hydrologique s'est modifié avec un apport d'eau douce résultant de l'abaissement du niveau des océans à la fin du Messinien et le déversement des grands fleuves d'Europe et d'Afrique (Èbre, Rhône, Pô et Nil) dans la dépression Méditerranéenne, formant localement des environnements lacustres. Le soubassement de la Méditerranée se situait localement à plus de 1000 mètres sous le niveau de l'océan Atlantique (par comparaison, aujourd'hui le niveau de la mer Morte en Israël se situe à -403 m, le lac Asal près de Djibouti à -155 m et la Vallée de la Mort en Utah à -85.5 m). À cette époque, on pouvait donc venir d'Afrique en Europe à pied et vice versa via la Méditerranée. Nous avons des preuves que des animaux vivant en Afrique du Nord ont profité de cet assèchement pour migrer vers le nord et inversement pour les populations du sud de l'Europe en empruntant les passages humides comme aujourd'hui on peut traverser les zones lacustres de l'Okavongo ou du delta du Nil en traversant les marais à la saison sèche. Bien que l'aventure était hasardeuse, on peut imaginer que des groupes d'homininés ont emprunté les mêmes voies. En revanche, en dehors de ces zones humides où la survie était possible, le désert de Méditerranée était une zone hostile qui devait représenter une barrière sans doute infranchissable et même mortelle en raison de l'ardeur des rayons du Soleil dans une région de haute pression à l'image des conditions extrêmes en termes de chaleur et de sécheresse régnant aujourd'hui dans la Vallée de la Mort en Utah, dans les salars de Bolivie ou d'Argentine ou encore dans le désert du Sahara où la température dépasse parfois 55°C à l'ombre et où le sol exposé toute une journée au Soleil peut atteindre 90°C. Puis, il y a environ 5.3 millions d'années, une brèche se produisit dans le détroit de Gibraltar qui s'élargit rapidement provoquant l'inondation cataclysmique de toute la partie occidentale de la Méditerranée jusqu'au versent occidental de l'Italie et de la Sicile dont les montagnes firent temporairement barrage à la mer. Cet évènement est appelé la méga inondation du Zancléen. Le scénario de déssication de la Méditerranée fut proposé en 1973 par K.J.Hsü et ses collègues puis fut révisé et complété en 1982 par Jean-Marie Rouchy et fut récemment mis à jour sur base de nouvelles données géologiques. Dans une étude publiée en 2018 dans la revue "Nature" par Aaron Micallef de l'Université de Malte et ses collègues, en utilisant des profils sismiques et des données de forage obtenues au large de la Sicile orientale, les chercheurs ont identifié un vaste corps de sédiments enfouis dans le sous-sol du canal de Sicile d'aspect chaotique. Ils ont nommé cette masse de matériel "Unité 2" qui est présentée ci-dessous à gauche. L’étude indique que ces énormes dépôts sédimentaires sont composés de matériaux érodés et transportés par un puissant flux d’eau qui inonda le bassin ionien par le détroit de Sicile après que le bassin occidental de la Méditerranée ait été rempli par l'eau de l’Atlantique.
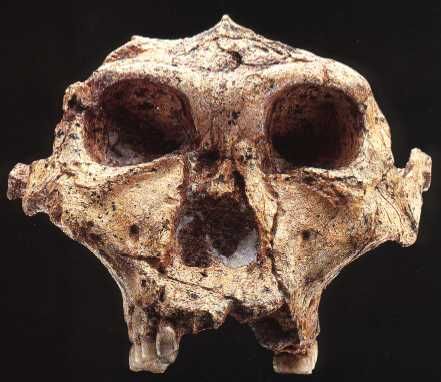
Comme le montre la coupe stratigraphique présentée ci-dessus à droite, les sédiments découverts (Unité 2 en brun) sont situés sur une couche de sels messiniens ainsi que sous une autre couche de sédiments marins déposés après la méga inondation, lorsque les conditions marines sont revenues à la normale. Selon les chercheurs, les dépôts identifiés sont sismiquement transparents et présentent une structure interne en couches désordonnées qui est très similaire aux sédiments qu'on observe généralement après des inondations catastrophiques (débacles ou crues). L'étude indique que la masse sédimentaire trouvée à proximité de la base de l'escarpement de Malte, entre l'est et l'ouest de la Méditerranée, forme un coin dont l'épaisseur a été estimée à 860 mètres tout près de l'escarpement. A ce jour, il s'agit de la plus grande trace de méga inondation connue sur Terre. Pour expliquer la présence de cette masse sédimentaire, les simulations (cf. la deuxième vidéo ci-dessus) montrent qu'à l'est de la Sicile, à hauteur du cañon de Noto se serait formée une cascade d'eau de 1.5 km de hauteur qui se déversa dans le cañon large de 5 km et déposa ses sédiments sur une longueur de 150 km où ils présentent encore une épaisseur de 12-13 mètres. L'eau noya rapidement la partie orientale du bassin méditerranéen puis encercla la Sicile avant d'envahir la future la mer Egée, la mer Adriatique et de grignoter une bonne partie des terres aujourd'hui inondées au large de l'actuelle Lybie et de la Tunisie. La mer Méditerranée présentait alors sa physionomie actuelle et forma une barrière qu'il était dorénavant impossible de franchir à pied et qu'il fallait contourner par le Moyen-Orient. Les chercheurs concluent que la méga inondation du Zancléen n’aurait duré que quelques années, le débit atteignant 100 millions de mètres cubes par seconde soit un taux ~1000 fois supérieur au débit actuel de l’Amazone ! Les chercheurs estiment qu'en deux ans seulement la Méditerranée s'est remplie d'eau jusqu'au niveau actuel ![2]. Sahelanthropus tchadensis alias Toumaï : 7 millions d'années Toumaï, qui signifie "espoir de vie" en langue Goran, fut découvert dans le désert du Djourab, au nord du Tchad, le 19 juillet 2001 par Ahounta Djimdoumalbaye, un membre de la Mission Paléoanthropologique Franco-Tchadienne (MPFT) dirigée par Michel Brunet du laboratoire PALEVOPRIM de l'Université de Poitiers. Djimdoumalbaye fut attiré par une pierre noire très irrégulière qui affleurait sur le sable du Sahel Tchadien, au nord de l'Afrique. Voici son commentaire : "je l'avais pris par l'arrière et je ne reconnus pas immédiatement l'intriguant objet. C'est après l'avoir retourné et encore retourné que je vis les cavités orbitales." C'est alors dit-il avec un large sourire que "j'ai senti mon rythme cardiaque s'accélérer. C'était un crâne fossilisé !" Djimdoumalbaye est déjà le découvreur de plusieurs autres fossiles d'hominidés.
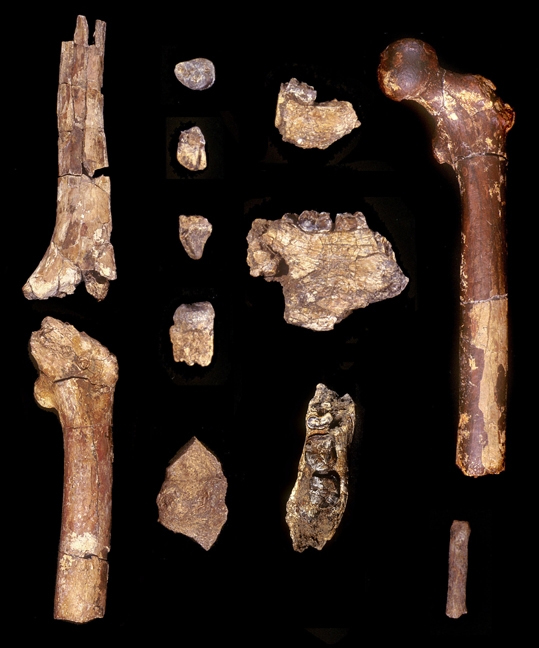
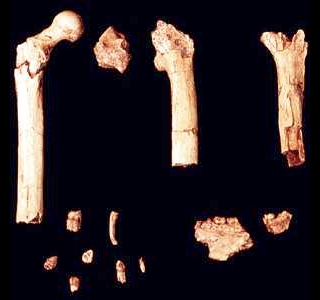
Le squelette de Toumaï fut découvert dans la région fossilifère TM 266 de Toros-Ménalla, au bord d'un paléo lac et est très incomplet. L'holotype est uniquement constitué d'un crâne subcomplet mais déformé. En l'espace de 5 ans, les paléoanthropologues ont découvert un seul crâne complet, plusieurs mâchoires inférieures, quelques dents isolées, la diaphyse (partie médiane) d'un fémur et des ulnae (cubitus), l'ensemble appartenant à quelque 9 individus apparentés à Toumaï (cf. "Nature", 418, 11 juillet 2002; 419, 10 octobre 2010; 434, 7 avril 2005). Après analyse, il s'avère que Toumaï compte parmi les plus vieux ancêtres de l'humanité et est le premier homininé découvert à l'ouest de la vallée du Rift. D'après le contexte géologique et le degré évolutif des fossiles de mammifères découverts sur le site, il est âgé d'environ 7 millions d'années. Il est né 350 000 générations avant nous ! Il appartient à une nouvelle espèce, Sahelanthropus tchadensis. Rappelons que le Tchad a déjà fait l'objet de plusieurs découvertes paléoanthropologiques. En 1959, Yves Coppens mit au jour le fossile d'un hominidé de l'espèce Tchadanthrope et l'équipe de Michel Brunet y découvrit l'Australopithèque Abel en 1995. Toumaï, Communiqué de Presse, CNRS, 2005
Quelles sont ses caractéristiques ? Son crâne disposant d'un important bourrelet sus orbitaire et d'attaches musculaires importantes, les paléoanthropologues estiment qu'il s'agit d'un mâle à la musculation puissante. Et comme ils disent, s'il s'agit d'un femelle, mieux valait ne pas rencontrer un mâle énervé ! Par extrapolation, on estime que sa taille est comprise entre 1.10 m et 1.30 m pour un poids oscillant entre 25 et 35 kg. L'individu est âgé d'une trentaine d'années. Il présente un front fuyant, en fait pas de front du tout associé à une capsule cérébrale allongée et étroite. La capacité de sa boîte crânienne est de 360 à 370 cm3, équivalente à celle des chimpanzés actuels. La face de Toumaï est haute et peu prognathe dans sa partie inférieure (sub-nasale). Elle présente une faible dépression nasale le rapprochant des homininés. Il ne présente pas de crête sagitale et n'est donc pas apparenté aux grands singes (Pan), ce n'est donc ni un chimpanzé ni un gorille. L'analyse de sa dentition révèle que ses canines supérieures ont été usées par la pointe et ses dents ont broyé des aliments durs et tendres, différents de ceux des grands singes. La morphologie de ses prémolaires et molaires présente un émail plus épais que celui des chimpanzés mais moins épais que celui des Australopithèques. Toutefois ses molaires présentent encore trois racines. Sa face est également plus plate que celle des grands singes. A
voir : L'évolution
de l'humanité depuis Toumaï
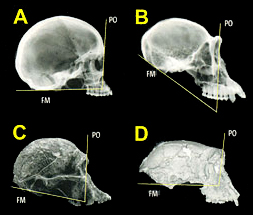
Enfin, la position du trou occipital et du Foramen magnum suggèrent que son crâne était fixé verticalement sur la colonne vertébrale et qu'il regardait donc droit devant lui. Ce fait est confirmé par l'angle supérieur à 90° formé par le plan de sa face nuquale (la nuque) très incliné vers l'arrière et le plan vertical des orbites, deux caractéristiques très proches de celles des Australopithèques et des Homo sapiens, ces deux seuls critères lui conférant le statut de bipède, qui plus est apparenté au genre préhumain. Toutefois, seule l'analyse de membres qui restent à découvrir pourra confirmer ou non sa posture verticale. L'affaire du "fémur de Toumaï" Notons que la diaphyse du fémur gauche que l'on voit sur la photo ci-dessous à gauche prise en 2001 ne fut analysée qu'en 2004 et la photo ne fut publiée dans la presse qu'en 2009 (cf. "La recherche", le site de John Hawks et l'article coécrit par Alain Beauvilain). En fait, comme le prouve l'autorisation d'exportation, le fémur ne fut pas expédié avec l'ensemble des fragments en 2001 mais s'est étrangement retrouvé dans la collection de Brunet en février 2004. L'analyse préliminaire effectuée par deux chercheurs extérieurs à la mission franco-tchadienne, Roberto Macchiarelli, auteur et paléoanthropologue de l'Université de Poitiers et du Muséum National d'Histoire Naturelle de Paris et Aude Bergeret, directrice du Musée d'Histoire Naturelle Victor-Brun de Montauban, confirma qu'il date d'environ 7 millions d'années mais malheureusement leurs résultats ne furent jamais publiés suite à l'embargo décrété par Brunet. Entre-temps Brunet prit sa retraite en 2005 puis le nouveau directeur du laboratoire PALEVOPRIM décréta unilatéralement un embargo sur le partage et la diffusion des informations avec l'accord tacite du CNRS alors que cette étude était pourtant attendue depuis des années par la communauté scientifique. Aussi l'appartenance de ce fémur à Toumaï fit l'objet d'une controverse dont la presse scientifique anglo-saxonne s'est faite l'écho tellement elle parut surréaliste. Pour éviter d'alourdir ce dossier, nous détaillerons cette affaire dans l'article consacré aux dérives de la science et en particulier à l'embargo et l'intolérance où nous décrirons également les suites de cette affaire. A lire : Le "fémur de Toumaï" ou le triste exemple de la dérive d'un directeur de recherche
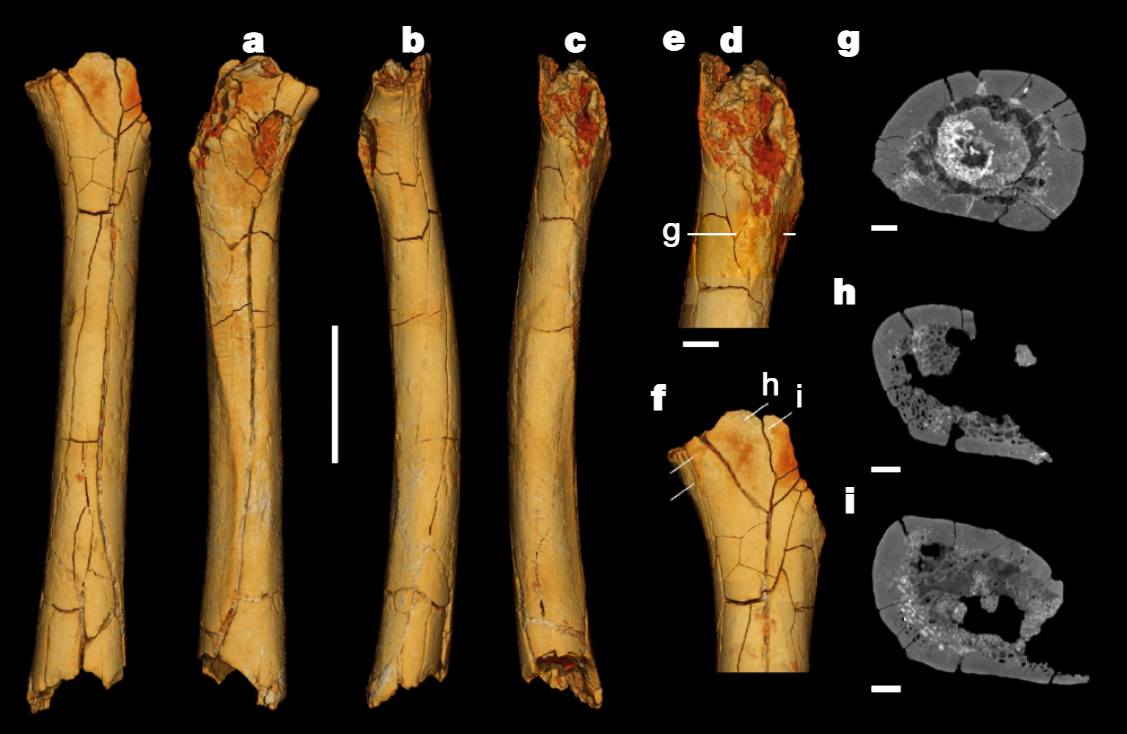
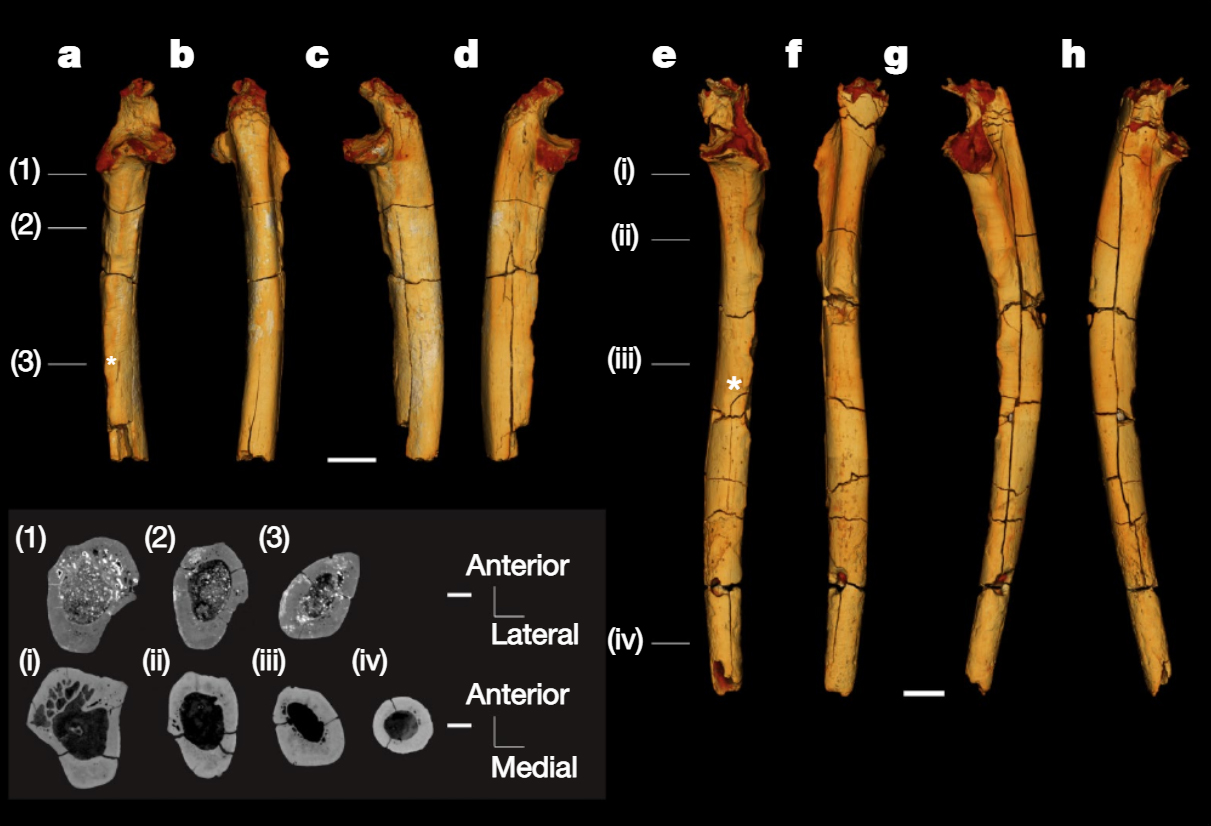
Finalement la diaphyse du fémur gauche et les deux ulnae (cubitus) droit et gauche du squelette de Toumaï découverts à la localité TM 266 furent analysés et firent l'objet d'une étude publiée dans la revue "Nature" (en PDF) en 2022 par l'équipe de Franck Guy du PALEVOPRIM de l'Université de Poitiers. Notons que Roberto Macchiarelli et Aude Bergeret précités qui participèrent à la première analyse n'ont pas cosigné cet article. Rien qu'en observant les ulnae (cubitus), on constate que l'os est très courbé par rapport à un ulna humain. Pour les paléoanthropologues, leur forme ressemble à celle de l'ulna de Lucy (voir page suivante). Il ne fait aucun doute qu'il ressemble aux ulnae des chimpanzés qui mènent une vie arboricole. Malheureusement on ne possède pas les os du pied de Toumaï qui pourraient appuyer cette hypothèse (comme on a pu le faire pour Ardipithecus ramidus, voir plus bas). Toutefois, l'obliquité du fémur qui se marque avec l'âge apporte un autre indice sur le fait que Toumaï était bipède. Les chercheurs sont arrivés à plusieurs conclusions. Ils constatent que si Toumaï était bipède il était aussi habile dans les arbres : "Nous présentons des preuves post-crâniennes du comportement locomoteur de S. tchadensis, avec de nouvelles connaissances sur la bipédie au stade précoce de l'histoire évolutive des homininés. [...] La morphologie du fémur est la plus parcimonieuse avec une bipédie habituelle, et les cubitus conservent la preuve d'un comportement arboricole important. Pris ensemble, ces résultats suggèrent que les hominidés étaient déjà bipèdes vers 7 Ma, mais suggèrent également que l'escalade arboricole était probablement une partie importante de leur répertoire locomoteur."
Selon Guy et ses collèguent, "le fémur de Sahelanthropus soutient les premières preuves de bipédie habituelle dans le clade des hominidés, confirmant ainsi les interprétations antérieures du matériel crânien basées sur l'orientation relative du plan orbitaire et du foramen magnum, ainsi que sur l'orientation et la morphologie du plan nucal. De plus, nos résultats post-crâniens favorisent une composante terrestre pour la bipédie habituelle chez S. tchadensis, comme le montrent O. tugenensis et A. kadabba tardif. Dans l'ensemble, les analyses des restes post-crâniens du Tchad suggèrent que l'adaptation à la bipédie a évolué peu après la divergence homme-chimpanzé, en conjonction avec le maintien d'adaptations ostéologiques pour les comportements positionnels arboricoles." Bien qu'il soit très primitif, Toumaï est sur la voie de l'hominisation; c'est bien notre ancêtre. La difficulté est de savoir où le placer dans l'arbre phylogénique car les restes des différentes espèces que nous possédons à l'heure actuelle sont très fragmentaires et ne facilitent pas les comparaisons. Toumaï est antérieur à Orrorin et Ardipithecus (voir plus bas) mais il s'en démarque suffisamment pour être placé à part. Pour prendre une analogie, nous dirions qu'il se place sur la partie droite de l'arborescence conduisant au genre Homo plutôt qu'à gauche. Contexte environnemental Concernant le biotope, l'analyse de la faune et de la flore fossiles situées dans les mêmes strates a permis de préciser que Toumaï vivait dans une région de paysages lacustres, avec des lacs, une faune aquatique, des espaces de savane arborée parsemée de prairies à graminées et de zones désertiques. On y trouvait une faune abondante (plus de 50% de bovidés et plus de 25% d'amphibiens) et très diversifiée, faisant de cet endroit un biotope inhabituel. On retrouve aujourd'hui une région analogue bien que moins étendue au Botswana, dans le delta de l'Okavango, dans le Kalahari central. Il est difficile d'imaginer que la présence de Toumaï en Afrique du Nord corresponde à une simple migration d'un groupe originaire du Rift vu que cette espèce existait plusieurs millions d'années avant toutes les autres. La découverte de Toumaï signifie que l'origine est-africaine de l'humanité (en Ethiopie) souvent évoquée par Yves Coppens et ses collègues doit être, sinon abandonnée, du moins révisée et élargie d'au moins... 2500 km. Nous verrons d'ici quelques années si les paléoanthropologues trouvent de nouveaux fossiles apparentés à cette espèce ainsi que l'éventuel "chaînon manquant" toujours absent de notre arbre généalogique et de quelle manière ils pourront décrire notre évolution en Afrique subtropicale.
Franck Guy et ses collègues soulignent également que "le matériel post-crânien de Toros-Ménalla ajoute aux interprétations précédentes du contexte environnemental des hominidés du Miocène du Miocène d'Afrique orientale connus jusqu'à présent. O. tugenensis est associé à des zones boisées ouvertes avec un couvert arboré important, Ardipithecus kadabba (dont la bipédie est déduite de la phalange plantaire AME-VP-1/71 datée de 5.2 Ma) est associée à un mélange de bois combinés avec des prairies humides, tandis qu'A. ramidus (4.4 Ma, Aramis, Éthiopie) habitait très probablement une forêt herbeuse alimentée par les eaux souterraines, probablement une palmeraie." "Pris dans leur ensemble, les premiers hominidés d'Afrique de l'Est partageaient une composante arboricole dans leur environnement. En ce qui concerne la zone fossilifère de Toros-Ménalla, les proxies paléoenvironnementaux suggèrent un paysage hétérogène qui comprend des forêts fermées (probablement des forêts riveraines), des palmeraies et des prairies mixtes (des forêts aux savanes et aux prairies aquatiques)." "Compte tenu des environs de Toros-Ménalla et du répertoire locomoteur inféré de S.tchadensis, les hominidés du Tchad ont pu exploiter à la fois les substrats arboricoles et terrestres pour se nourrir et accéder aux ressources en eau. L'association entre un répertoire locomoteur diversifié (dans les arbres et au sol) et des habitats boisés en contexte mésique depuis au moins environ 2.5 millions d'années suggère que la niche écologique de ces premiers hominidés n'était pas nécessairement liée à l'expansion de zones relativement sèches et ouvertes. Cette niche pourrait être décrite comme opportuniste dans sa dépendance à l'égard des ressources terrestres et arboricoles." "Sur la base de données moléculaires, on a estimé que le dernier ancêtre commun chimpanzé-humain se produisait en Afrique entre 10 et 6 Ma. Les représentants fossiles du clade des paninés sont rares, mais au moins trois taxons d'hominidés ont été décrits dans cet intervalle de temps en Afrique : Samburupithecus, de Samburu Hills, vers 9.5 Ma; Nakalipithecus de Nakali, vers 9.8 Ma ; et Chororapithecus de Chorora, il y a environ 8 Ma. Ces taxons miocènes sont parcimonieusement attribués aux hominines souches, même si Samburupithecus présente une morphologie particulièrement archaïque. Chororapithecus affiche des affinités dentaires dérivées avec Gorilla. À la lumière de cet enregistrement et du manque de résolution phylogénétique, la condition ancestrale du comportement positionnel chez les singes d'Afrique et les humains restera insaisissable jusqu'à ce que de nouvelles données significatives soient disponibles. À ce jour, l'identification des traits dérivés partagés par les homininés repose sur l'analyse des premiers taxons du clade." (cf. F.Guy et al., 2022). Orrorin tugenensis : 6 millions d'années A l’automne 2000, Brigitte Senut et Martin Pickford du Muséum National d'Histoire Naturelle de Paris et des membres de la Kenya Palaeontology Expedition (KPE) découvrirent dans la formation de Lukeino située près du lac Baringo, dans les collines Tugen à l'ouest du Kenya, quelques fragments d'os longs et de canines appartenant à la même espèce que la molaire inférieure découverte en 1974 à Lukeino et dont le propriétaire n'était pas clairement identifié (hominidé ou chimpanzé, les données étaient insuffisantes pour le préciser). Des fossiles appartenant à la même espèce furent également découverts dans quatre sites proches situés dans la partie sud du bassin Baringo, dans une zone de 50x30 km : à Aragai, Cheboit, Kapcheberek et Kapsomin. Yves Coppens assura la publication dans les "Comptes Rendus de l'Académie des Sciences" en janvier 2001. Après quelques hésitations sur le propriétaire des dents, les fossiles furent crédités à un préhumain bipède mais rattaché à un nouverau taxon, le "Millenium Ancestor" (Ancêtre du Millénaire), Orrorin tugenensis. Son squelette daterait entre 6.1 et 5.7 millions d'années. C'est à Kapsomin qu'on récolta le plus grand nombre de fossiles. Malheureusement aucun fragment de crâne ne fut découvert. Le gisement est situé près d'une falaise basse de basalte en bordure du lac. De nombreux fragments d’Orrorin ayant été trouvés au pied de la falaise, les paléoanthropologues estiment qu'elle fut la proie d'un carnivore, probablement un félin proche d’un léopard dont les restes furent découverts sur le site.
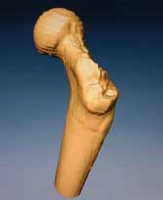
La dimension de ses os suggère qu'Orrorin mesurait entre 1.25 m et 1.50 m et pesait entre 30 et 45 kg. Par sa morphologie dentaire, Orrorin présente des caractères à la fois archaïques et modernes. D'un côté ses dents sont de taille modeste, bien inférieure à celle des Australopithèques, d'un autre côté elles sont proches de celles des chimpanzés et des hommes. Les dents découvertes à Kapsomin éloignent Orrorin des grands singes par plusieurs caractères (absence de forte crénulation aux molaires, un épaulement de la couronne de la canine inférieure, émail épaissi aux molaires, etc). Parmi les caractères homininés, on peut noter l’absence d'espace (diastème) à la mâchoire inférieure et la présence d’un tubercule distal à la canine inférieure. L’étude des os longs d’Orrorin suggère qu'elle était bipède. En effet, son fémur ne présente pas de caractères dérivés des grands singes mais il rappelle ceux des Australopithèques et de l’homme. Ils sont donc primitifs (à l'image des os de l'Ugandapithecus, du Proconsul ou du Kenyapithecus africanus). La tête fémorale est également plus grande proportionnellement que le col et tordue vers l’avant; deux morphologies qui rappellent celles de l’homme. Chez les Australopithèques, le diamètre de la tête fémorale est seulement un peu plus grand que celui du col et la tête est orientée médialement à postérieurement. Il ressort de toutes ces données que non seulement Orrorin marchait sur ses deux pieds, mais se dandinait différemment des Australopithèques, tout en se rapprochant de la posture de l’homme. Enfin, les restes postcrâniens suggèrent des adaptations arboricoles tandis que la forme de sa première phalange indique qu'elle avait la faculté de se suspendre et de grimper aux arbres (humérus aplati, courbure et longueur de la phalange proximale).
Où placer Orrorin dans l'arbre phylogénique ? Ses os sont plus grands que ceux de Lucy, mais ses molaires sont plus petites (microdontie), la rapprochant des caractères des préhumains du Plio-Pléistocène et des hommes modernes. Sa présence chez un homininé d'environ 6 millions d'années suggère que son espèce était déjà présente avant cette époque. La microdontie indique un régime riche peut-être frugivore ou même omnivore et non plus végétarien (mégadontie). Ces caractères semblent indiquer que la divergence des homininés bipèdes des grands singes africains se situerait entre 9 et 8 millions d'années, date compatible avec la découverte de Toumaï à 7 millions d'années. Ce chaînon manquant serait apparu juste après Samburupithecus qui habita dans les savanes d’Afrique orientale il y a 9.5 millions d'années. La découverte d'Orrorin suggère également que la séparation des Australopithèques du genre Homo s’est bien produite plutôt qu’on le pensait, probablement vers 8 millions d'années, période qui a connu un changement important dans les communautés mammaliennes africaines.
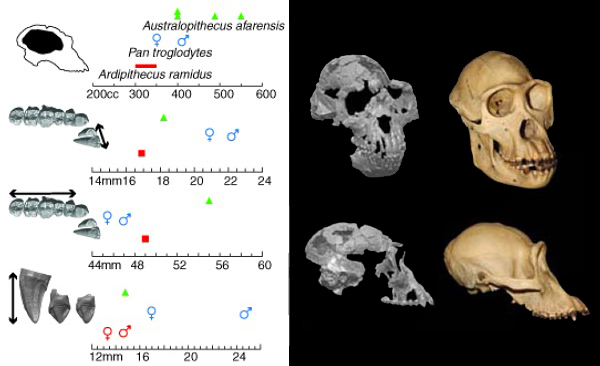
Orrorin est un maillon très important de notre arbre phylogénique. Il représente une espèce hybride, mélangeant des caractères humains et simiesques. Comme Toumaï, il présente une face assez plate, une posture verticale et un régime alimentaire différents des grands singes plusieurs millions d'années avant Abel et Lucy. Orrorin est paradoxalement plus humaine que Lucy qui pourtant est 3 millions d’années plus jeune qu'elle. On peut donc suggérer sans commettre trop d'erreurs que les Australopithèques ne sont pas nos ancêtres directs, mais représentent une branche latérale de notre arbre généalogique. Comme évoqué plus haut, en 2002 on découvrit en Crète des empreintes dans des sédiments datant de 5.7 millions d'années. Ils furent attribués à des homininés et non pas à des hominidés et encore moins à des singes. Le sujet touchant la station debout des premiers préhumains, on y reviendra à propos de la bipédie. Ardipithecus ramidus : 4.4 millions d'années C'est pendant une mission en Ethiopie qui eut lieu entre 1992 et 1993 puis en 1994 que l'équipe du professeur Tim White de l’Université de Californie découvrit dans les strates sédimentaires du Pliocène 125 fragments d’os et des dents provenant de 36 individus appartenant à une nouvelle espèce d'homininés qui, après hésitation, sera appelée Ardipithecus ramidus. Cette espèce vivait il y a 4.4 millions d'années. La découverte fut l'objet d'une publication dans la revue "Nature".
L'un des squelettes les mieux étudiés (ARA-VP-6/500) correspond à une personne mesurant 1.20 m et pesant environ 50 kg, des mensurations assez importantes pour l'époque si on les compare à la petite Lucy. En analysant ces squelettes et les fossiles de la flore de cette époque, Tim White et son équipe firent des découvertes fondamentales sur la manière dont cet homininé se déplaçait, comment il marchait et accessoirement dans quel biotope il vivait. En effet, la première découverte est le fait qu'Ardipithecus ramidus présente des mains dont le pouce est opposable. Cette capacité est plutôt précoce car encore aujourd'hui si la plupart des singes (chimpanzé, bonobo, orang-outan, singe écureuil, etc) présentent cette particularité, d'autres espèces comme le ouistiti vivant dans le Nouveau Monde ne l'a pas acquise. De plus les os des phalanges des mains d'A.ramidus ne sont pas usées comme chez les singes posant le dos de leur main au sol pour marcher, indice supplémentaire que A.ramidus se déplaçait différemment des autres singes. En revanche, comme tous les singes, A.ramidus présente des pieds dont le gros orteil est opposable, confirmant son adaptation à la vie arboricole, le pouce et le gros orteil opposables lui permettant de s'agripper aux branches les plus fines, celles portant les fleurs et les fruits.
Plus étonnant, le pelvis c'est-à-dire les os du bassin (os sacrum, coccyx et les deux os iliaques) d'A.ramidus supporte l'hypothèse que cette espèce était capable de se tenir debout naturellement sans être déhanchée. En effet, le haut de son bassin ressemble au nôtre, mais le bas est plus haut que celui des humains; il est mieux adapté à la vie arboricole. Il pouvait donc marcher sur ses deux jambes mais pas aussi bien que Lucy et des autres homininés apparus plus tard. A partir de la tailles des membres et des dents des mâles et des femelles A.ramidus, on a également découvert qu'il y avait peu de dimorphisme sexuel, une caractéristique également précoce qui apparaît surtout chez les grands singes et le genre Homo. Comme chez les bonobos, cela peut aussi signifier qu'ils vivaient dans un environnement stable où il n'y avait pas de rivalités entre mâles ou entre espèces. Enfin, l'analyse biochronologique des strates révèle qu'entre 40 et 65% des centaines d'espèces végétales répertoriées se développaient dans une zone forestière et non pas dans la savane. Cela suggère qu'Ardipithecus ramidus vivait en forêt. Vu l'adaptation de ses pieds, il était donc vraisemblablement arboricole, aussi à l'aise dans les arbres que nos chimpanzés tout en pouvant profiter de la bipédie pour se redresser et marcher au sol.
Grâce à des méthodes comparatives de morphométrie et de phylogénétique, des chercheurs ont montré qu'Ardipithecus conserva dans ses mains des morphologies adaptées à la suspension partagées avec les chimpanzés et les bonobos. Dans l'ensemble, ces résultats suggèrent que les premiers hominidés ont évolué à partir d'un ancêtre ayant un répertoire de position varié comprenant la suspension et l'escalade verticale, affectant directement les hypothèses sur l'origine de la lignée humaine (cf. T.Prang et al., 2021). A partir de ces caractéristiques hybrides entre celles des singes et des préhumains, à ce jour un cas unique dans l'histoire de l'humanité, cette espèce fut qualifiée de "ramidus" pour préciser que cet homininé est situé à la racine de la lignée qui conduit au genre Homo. Cet homininé archaïque bipède et présentant un petit cerveau vivait 500000 ans avant Lucy, considérée comme le plus ancien fossile d'Australopithecus afarensis. Prochain chapitre
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||




































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}